

Structural studies of the actions of anesthetic drugs on the γ-aminobutyric acid type A receptor
- PMID: 21881491
- PMCID: PMC3226907
- DOI: 10.1097/ALN.0b013e3182315d93
Structural studies of the actions of anesthetic drugs on the γ-aminobutyric acid type A receptor
Abstract
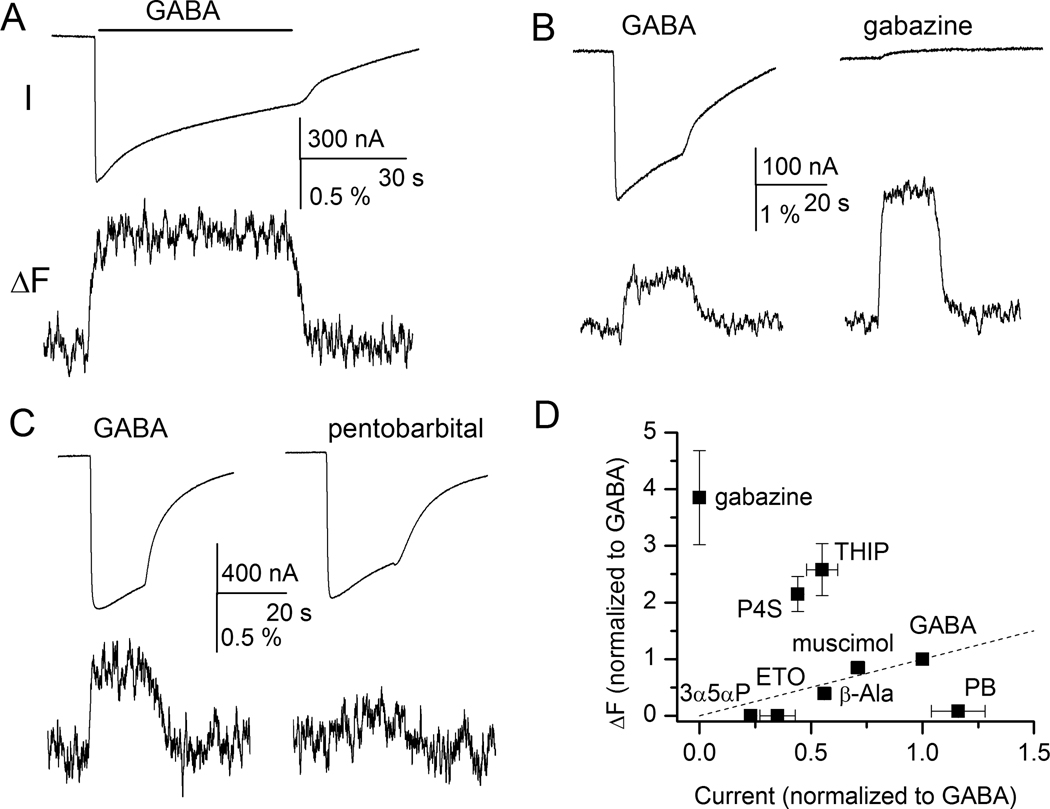
The γ-aminobutyric acid type A receptor is the major transmitter-gated inhibitory channel in the central nervous system. The receptor is a target for anesthetics, anticonvulsants, anxiolytics, and sedatives whose actions facilitate the flow of chloride ions through the channel and enhance the inhibitory tone in the brain. Both the kinetic and structural aspects of the actions of modulators of the γ-aminobutyric acid type A receptor are of great importance to understanding the molecular mechanisms of general anesthesia. In this review, the structural rearrangements that take place in the γ-aminobutyric acid type A receptor during channel activation and modulation are described, focusing on data obtained using voltage-clamp fluorometry. Voltage-clamp fluorometry entails the binding of an environmentally sensitive fluorophore molecule to a site of interest in the receptor, and measurement of changes in the fluorescence signal resulting from activation- or modulation-elicited structural changes. Detailed investigations can provide a map of structural changes that underlie or accompany the functional effects of modulators.
Figures



References
-
- Bonin RP, Orser BA. GABAA receptor subtypes underlying general anesthesia. Pharmacol Biochem Behav. 2008;90:105–112. - PubMed
-
- Franks NP. General anaesthesia: From molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9:370–386. - PubMed
-
- Jones-Davis DM, Macdonald RL. GABAA receptor function and pharmacology in epilepsy and status epilepticus. Curr Opin Pharmacol. 2003;3:12–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
