

13C MRS studies of neuroenergetics and neurotransmitter cycling in humans
- PMID: 21882281
- PMCID: PMC3651027
- DOI: 10.1002/nbm.1772
13C MRS studies of neuroenergetics and neurotransmitter cycling in humans
Abstract
In the last 25 years, (13)C MRS has been established as the only noninvasive method for the measurement of glutamate neurotransmission and cell-specific neuroenergetics. Although technically and experimentally challenging, (13)C MRS has already provided important new information on the relationship between neuroenergetics and neuronal function, the energy cost of brain function, the high neuronal activity in the resting brain state and how neuroenergetics and neurotransmitter cycling are altered in neurological and psychiatric disease. In this article, the current state of (13)C MRS as it is applied to the study of neuroenergetics and neurotransmitter cycling in humans is reviewed. The focus is predominantly on recent findings in humans regarding metabolic pathways, applications to clinical research and the technical status of the method. Results from in vivo (13)C MRS studies in animals are discussed from the standpoint of the validation of MRS measurements of neuroenergetics and neurotransmitter cycling, and where they have helped to identify key questions to address in human research. Controversies concerning the relationship between neuroenergetics and neurotransmitter cycling and factors having an impact on the accurate determination of fluxes through mathematical modeling are addressed. We further touch upon different (13)C-labeled substrates used to study brain metabolism, before reviewing a number of human brain diseases investigated using (13)C MRS. Future technological developments are discussed that will help to overcome the limitations of (13)C MRS, with special attention given to recent developments in hyperpolarized (13)C MRS.
Copyright © 2011 John Wiley & Sons, Ltd.
Figures

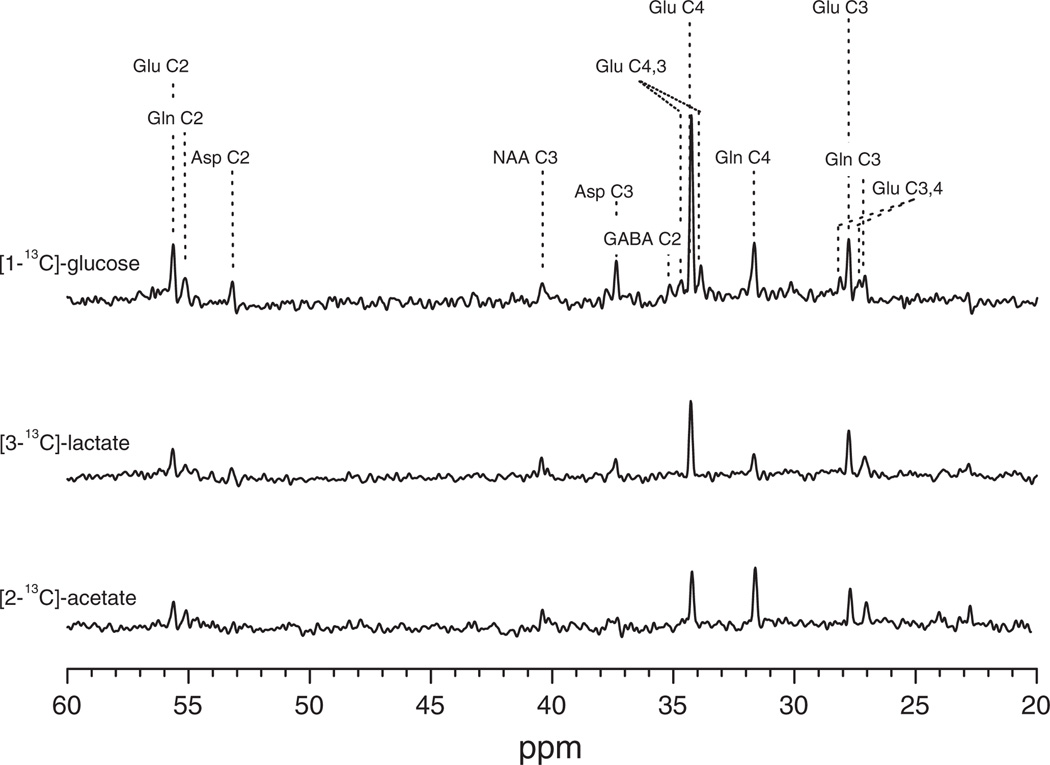

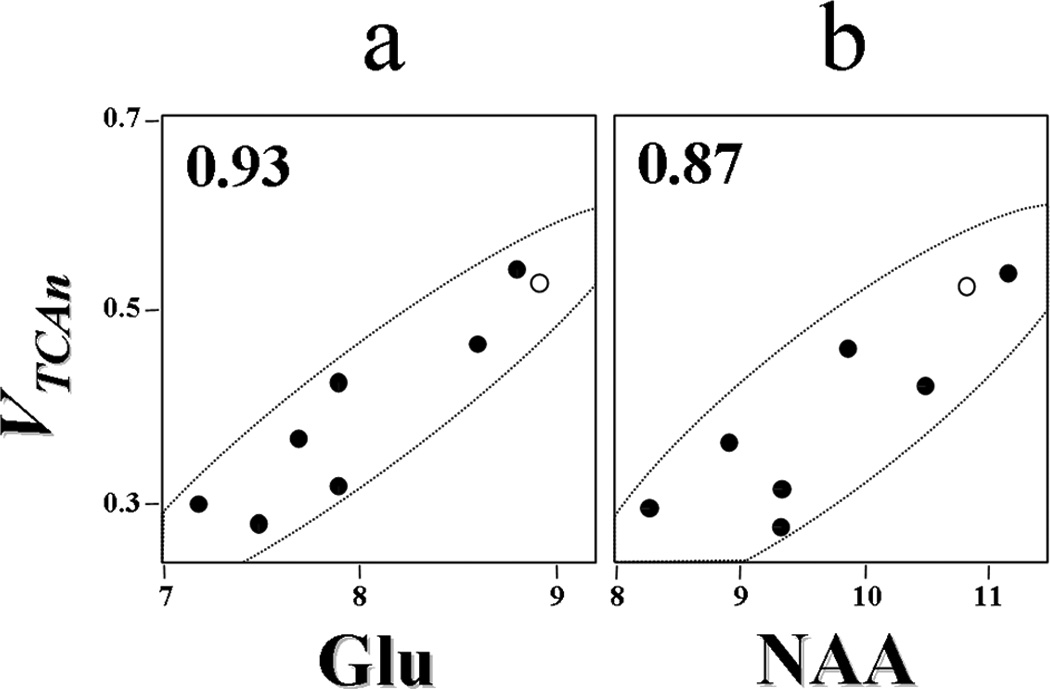

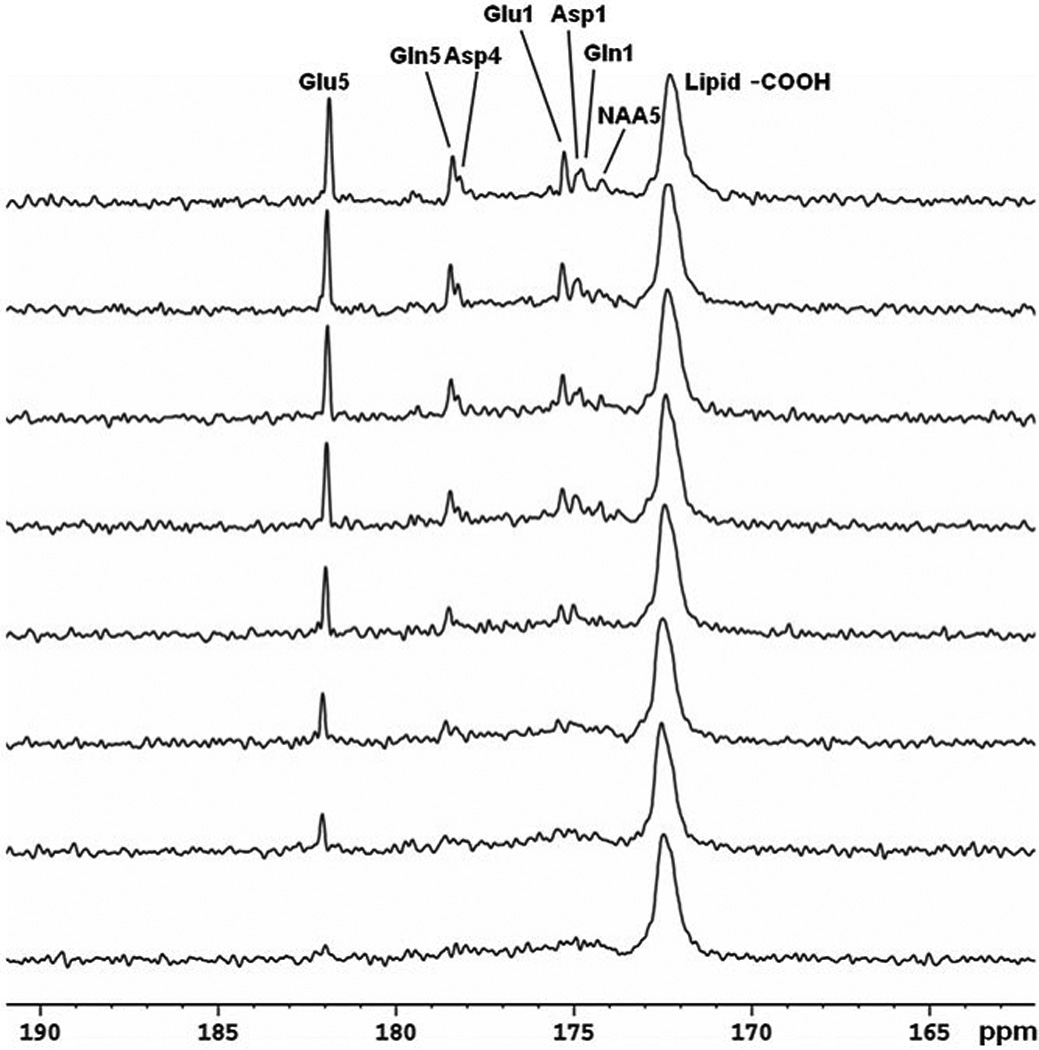
References
-
- Shephard GM. The synaptic organization of the brain. Oxford, U.K: Oxford University Press; 1994.
-
- Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, Kanai Y, Hediger MA, Wang Y, Schielke JP, Welty DF. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16(3):675–686. - PubMed
-
- Bergles DE, Diamond JS, Jahr CE. Clearance of glutamate inside the synapse and beyond. Curr Opin Neurobiol. 1999;9(3):293–298. - PubMed
-
- Hertz L, Peng L, Dienel GA. Energy metabolism in astrocytes: high rate of oxidative metabolism and spatiotemporal dependence on glycolysis/glycogenolysis. J Cereb Blood Flow Metab. 2007;27(2):219–249. - PubMed
-
- Sibson NR, Mason GF, Shen J, Cline GW, Herskovits AZ, Wall JE, Behar KL, Rothman DL, Shulman RG. In vivo (13)C NMR measurement of neurotransmitter glutamate cycling, anaplerosis and TCA cycle flux in rat brain during. J Neurochem. 2001;76(4):975–989. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-DA021785/DA/NIDA NIH HHS/United States
- P30-NS052519/NS/NINDS NIH HHS/United States
- R21 AA018210/AA/NIAAA NIH HHS/United States
- R01 DA021785/DA/NIDA NIH HHS/United States
- R41 EB008296/EB/NIBIB NIH HHS/United States
- R01-EB000473/EB/NIBIB NIH HHS/United States
- R21-AA019803/AA/NIAAA NIH HHS/United States
- R01 EB009336/EB/NIBIB NIH HHS/United States
- R01 MH095104/MH/NIMH NIH HHS/United States
- R21-AA018210/AA/NIAAA NIH HHS/United States
- R01-MH95104/MH/NIMH NIH HHS/United States
- R01 AG034953/AG/NIA NIH HHS/United States
- R01-DK027121/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
