

Review
doi: 10.1016/j.stem.2011.08.007.
TET family proteins and their role in stem cell differentiation and transformation
Affiliations
- PMID: 21885017
- PMCID: PMC3244690
- DOI: 10.1016/j.stem.2011.08.007
Item in Clipboard
Review
TET family proteins and their role in stem cell differentiation and transformation
Cell Stem Cell.
.
Abstract
One of the main regulators of gene expression during embryogenesis and stem cell differentiation is DNA methylation. The recent identification of hydroxymethylcytosine (5hmC) as a novel epigenetic mark sparked an intense effort to characterize its specialized enzymatic machinery and to understand the biological significance of 5hmC. The recent discovery of recurrent deletions and somatic mutations in the TET gene family, which includes proteins that can hydroxylate methylcytosine (5mC), in a large fraction of myeloid malignancies further suggested a key role for dynamic DNA methylation changes in the regulation of stem cell differentiation and transformation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
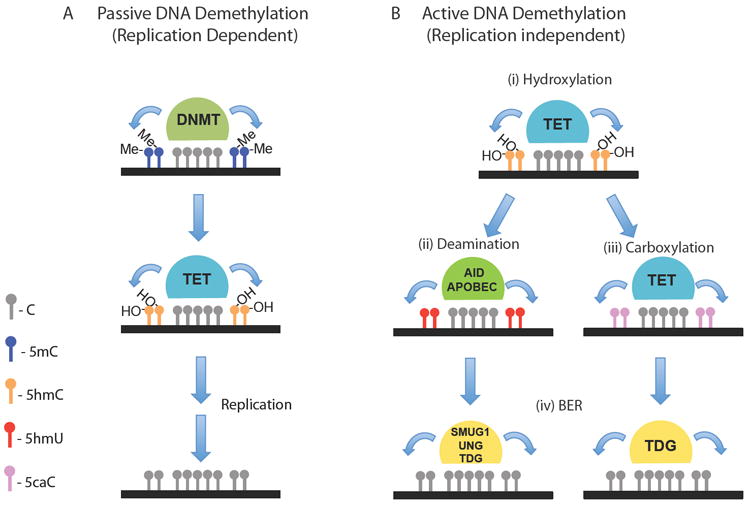
A) The inability of the maintenance methyltransferase, DNMT1, to recognize 5hmC-containing DNA may lead to passive demethylation following replication. B) Active DNA methylation can be catalyzed by TET-mediated (i) hydroxylation of 5mC to 5hmC followed by AID and APOBEC-mediated (ii) deamination of 5hmC to 5hmU or (iii) further carboxylation of 5hmC to 5caC by the TET proteins and the subsequent cleavage of 5hmU or 5caC by DNA glycosylases (e.g. SMUG1, UNG and TDG) and replacement with cytosine by (iv) a base excision repair (BER) enzymatic pathway.
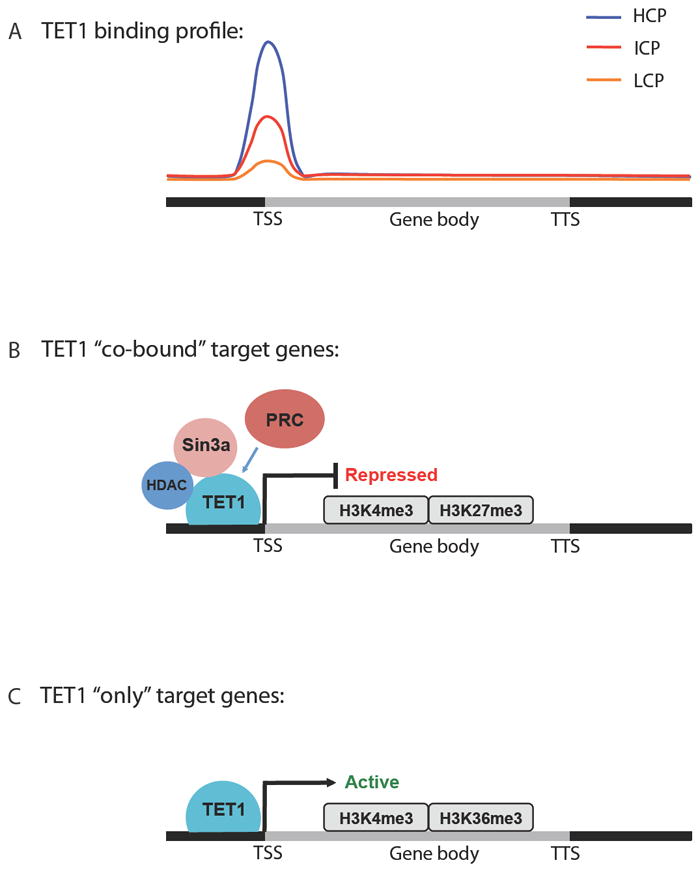
A) Schematic of TET1 binding profile in the genome of mouse ESCs. TET1 is most highly enriched at the transcriptional start site (TSS) of genes with high CpG promoters (HCPs) compared to those with intermediate and low CpG promoters (ICPs and LCPs, respectively). B) TET1 directly binds to the co-repressors Sin3a and HDACs. TET1 target genes co-bound by polycomb repressor complex (PRC) proteins and Sin3a are repressed and display bivalent H3K4me3 and H3K27me3 marks. C) TET1-only target genes are actively transcribed and enriched for H3K4me3 and H3K36me3.
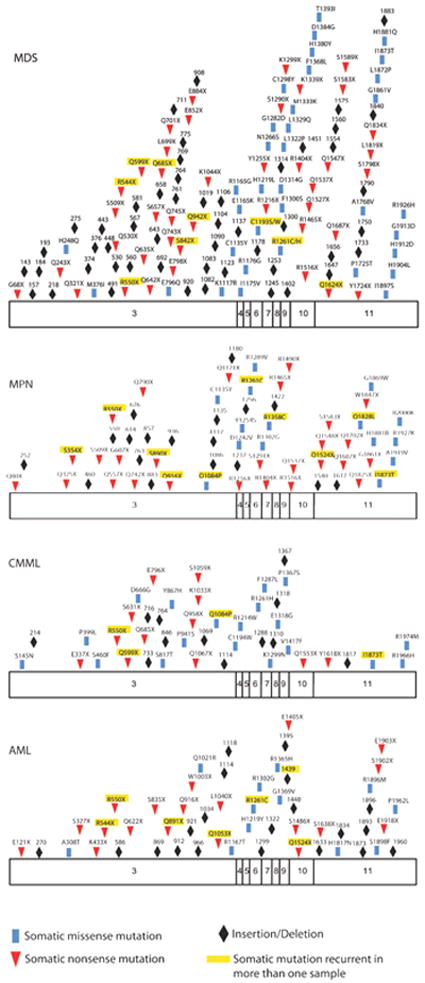
Displayed are the known somatic missense, nonsense, and frameshift mutations throughout the open-reading frame of TET2 in myelodisplastic syndromes (MDS), myeloproliferative neoplasms (MPN), chronic myelomonocytic leukemia (CMML) and acute myeloid leukemia (AML). In addition mutations that were seen in more than one sample amongst studies are highlighted in yellow. Data are compiled from several studies (Abdel-Wahab et al., 2009; Bejar et al., 2011; Delhommeau et al., 2009; Figueroa et al., 2010; Jankowska et al., 2009; Koh et al., 2011).
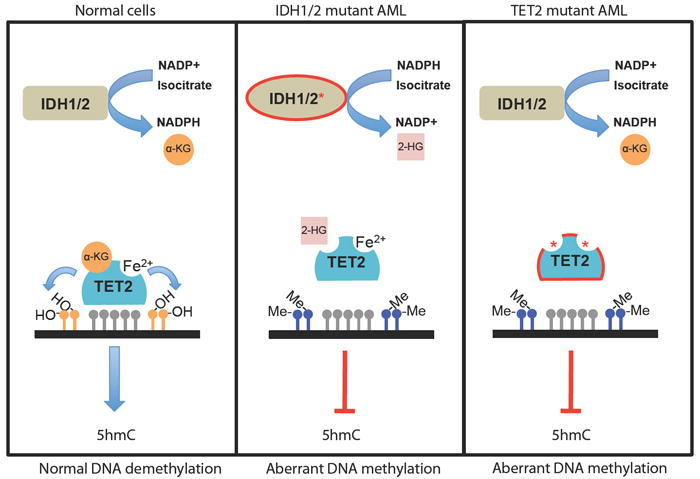
IDH proteins normally catalyze the conversion of NADP+ into NADPH and isocitrate into α-ketoglutarate (α-KG), a substrate required for normal TET2 enzymatic activity. Mutant IDH proteins cause loss of function of TET2 by exhibiting neomorphic enzymatic activity that results in the consumption of NADPH and production of the oncometabolite 2-hydroxyglutarate (2-HG) instead of α-KG. IDH mutations are mutually exclusive with TET2 mutations found in patients that exhibit reduced hydroxylase activity, due in part to mutations in the 2-OG and Fe(II)-dependant dioxygenase domain that cause decreased 5hmC production and aberrant DNA methylation patterns.
References
-
- Barreto G, Schafer A, Marhold J, Stach D, Swaminathan SK, Handa V, Doderlein G, Maltry N, Wu W, Lyko F, et al. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature. 2007;445:671–675. - PubMed
-
- Bhattacharya SK, Ramchandani S, Cervoni N, Szyf M. A mammalian protein with specific demethylase activity for mCpG DNA. Nature. 1999;397:579–583. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U54 CA143798/CA/NCI NIH HHS/United States
- R01 CA133379/CA/NCI NIH HHS/United States
- R01CA133379/CA/NCI NIH HHS/United States
- R21CA141399/CA/NCI NIH HHS/United States
- R01 CA149655/CA/NCI NIH HHS/United States
- R01 CA105129/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R21 CA141399/CA/NCI NIH HHS/United States
- R01CA149655/CA/NCI NIH HHS/United States
- R01CA105129/CA/NCI NIH HHS/United States
- 1R01CA138234-01/CA/NCI NIH HHS/United States
- R01GM088847/GM/NIGMS NIH HHS/United States
- R01 CA138234/CA/NCI NIH HHS/United States
- U54CA143798-01/CA/NCI NIH HHS/United States
- R01 GM088847/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
