

Genome-wide maps of histone modifications unwind in vivo chromatin states of the hair follicle lineage
- PMID: 21885018
- PMCID: PMC3166618
- DOI: 10.1016/j.stem.2011.07.015
Genome-wide maps of histone modifications unwind in vivo chromatin states of the hair follicle lineage
Abstract
Using mouse skin, where bountiful reservoirs of synchronized hair follicle stem cells (HF-SCs) fuel cycles of regeneration, we explore how adult SCs remodel chromatin in response to activating cues. By profiling global mRNA and chromatin changes in quiescent and activated HF-SCs and their committed, transit-amplifying (TA) progeny, we show that polycomb-group (PcG)-mediated H3K27-trimethylation features prominently in HF-lineage progression by mechanisms distinct from embryonic-SCs. In HF-SCs, PcG represses nonskin lineages and HF differentiation. In TA progeny, nonskin regulators remain PcG-repressed, HF-SC regulators acquire H3K27me3-marks, and HF-lineage regulators lose them. Interestingly, genes poised in embryonic stem cells, active in HF-SCs, and PcG-repressed in TA progeny encode not only key transcription factors, but also signaling regulators. We document their importance in balancing HF-SC quiescence, underscoring the power of chromatin mapping in dissecting SC behavior. Our findings explain how HF-SCs cycle through quiescent and activated states without losing stemness and define roles for PcG-mediated repression in governing a fate switch irreversibly.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
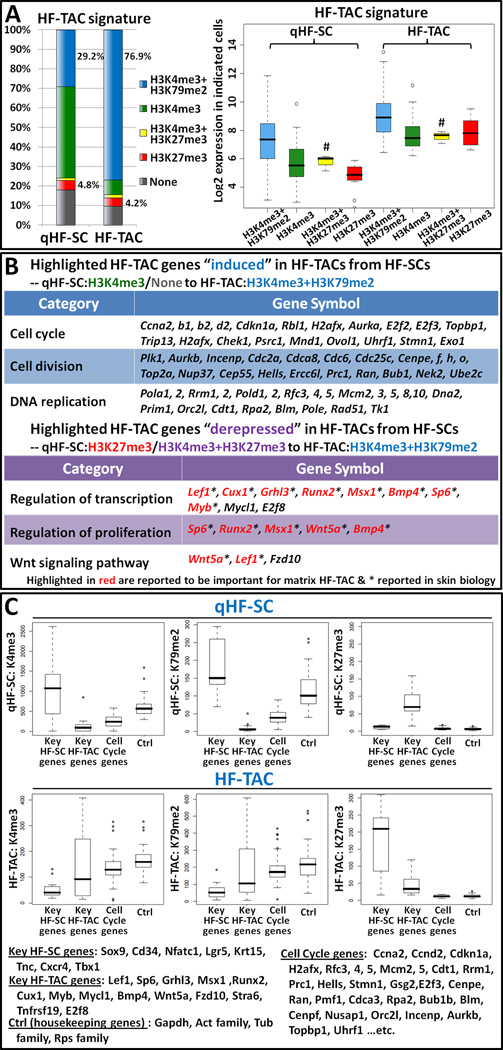
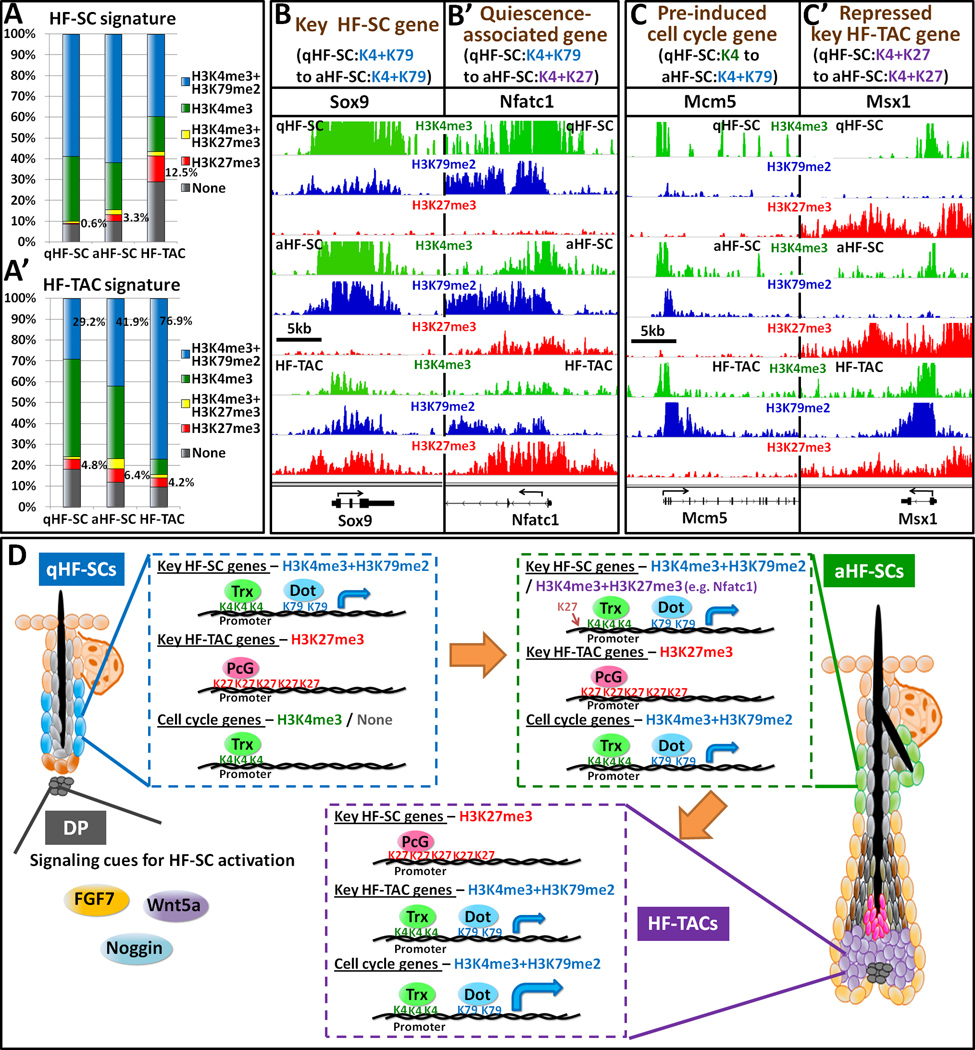

Comment in
-
Committing to a hairy fate: epigenetic regulation of hair follicle stem cells.Cell Stem Cell. 2011 Sep 2;9(3):183-4. doi: 10.1016/j.stem.2011.08.009. Cell Stem Cell. 2011. PMID: 21885014
References
-
- Bardot B, Lecoin L, Fliniaux I, Huillard E, Marx M, Viallet JP. Drm/Gremlin, a BMP antagonist, defines the interbud region during feather development. Int J Dev Biol. 2004;48:149–156. - PubMed
-
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, Jaenisch R, Wagschal A, Feil R, Schreiber SL, Lander ES. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125(2):315–326. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
