

Adipose-selective overexpression of ABHD5/CGI-58 does not increase lipolysis or protect against diet-induced obesity
- PMID: 21885429
- PMCID: PMC3196235
- DOI: 10.1194/jlr.M019117
Adipose-selective overexpression of ABHD5/CGI-58 does not increase lipolysis or protect against diet-induced obesity
Abstract
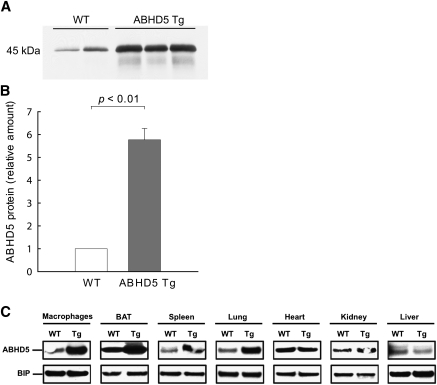
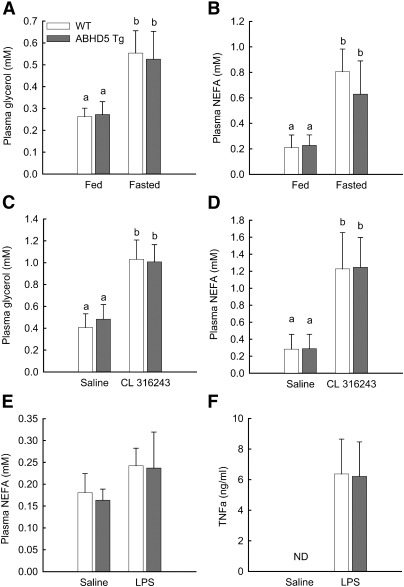
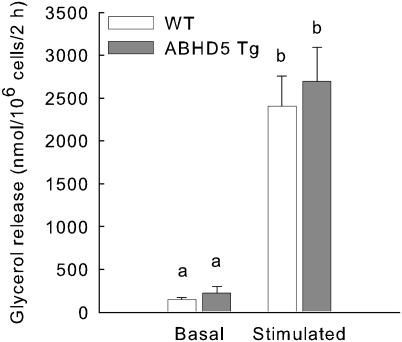
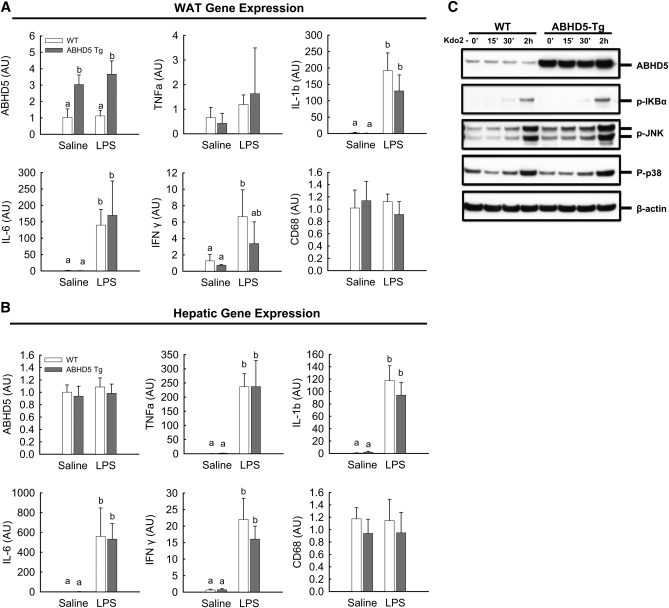
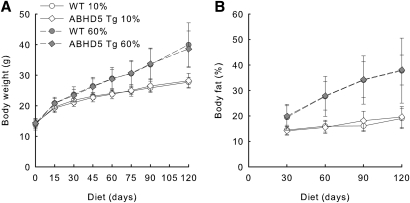
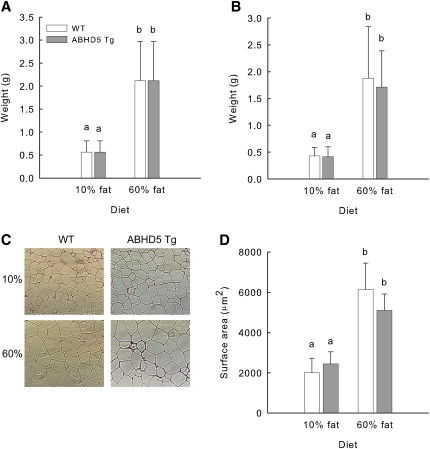
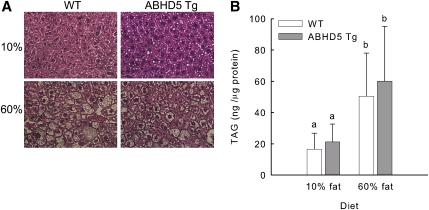
Adipose triglyceride lipase (ATGL) catalyzes the first step of triacylglycerol hydrolysis in adipocytes. Abhydrolase domain 5 (ABHD5) increases ATGL activity by an unknown mechanism. Prior studies have suggested that the expression of ABHD5 is limiting for lipolysis in adipocytes, as addition of recombinant ABHD5 increases in vitro TAG hydrolase activity of adipocyte lysates. To test this hypothesis in vivo, we generated transgenic mice that express 6-fold higher ABHD5 in adipose tissue relative to wild-type (WT) mice. In vivo lipolysis increased to a similar extent in ABHD5 transgenic and WT mice following an overnight fast or injection of either a β-adrenergic receptor agonist or lipopolysaccharide. Similarly, basal and β-adrenergic-stimulated lipolysis was comparable in adipocytes isolated from ABHD5 transgenic and WT mice. Although ABHD5 expression was elevated in thioglycolate-elicited macrophages from ABHD5 transgenic mice, Toll-like receptor 4 (TLR4) signaling was comparable in macrophages isolated from ABHD5 transgenic and WT mice. Overexpression of ABHD5 did not prevent the development of obesity in mice fed a high-fat diet, as shown by comparison of body weight, body fat percentage, and adipocyte hypertrophy of ABHD5 transgenic to WT mice. The expression of ABHD5 in mouse adipose tissue is not limiting for either basal or stimulated lipolysis.
Figures
References
-
- Langin D. 2006. Control of fatty acid and glycerol release in adipose tissue lipolysis. C. R. Biol. 329: 598–607 - PubMed
-
- Schweiger M., Schreiber R., Haemmerle G., Lass A., Fledelius C., Jacobsen P., Tornqvist H., Zechner R., Zimmermann R. 2006. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 281: 40236–40241 - PubMed
-
- Zechner R., Strauss J. G., Haemmerle G., Lass A., Zimmermann R. 2005. Lipolysis: pathway under construction. Curr. Opin. Lipidol. 16: 333–340 - PubMed
-
- Zimmermann R., Strauss J. G., Haemmerle G., Schoiswohl G., Birner-Gruenberger R., Riederer M., Lass A., Neuberger G., Eisenhaber F., Hermetter A., et al. 2004. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 306: 1383–1386 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
