

AMP-activated protein kinase enhances the phagocytic ability of macrophages and neutrophils
- PMID: 21885655
- PMCID: PMC3236633
- DOI: 10.1096/fj.11-190587
AMP-activated protein kinase enhances the phagocytic ability of macrophages and neutrophils
Abstract
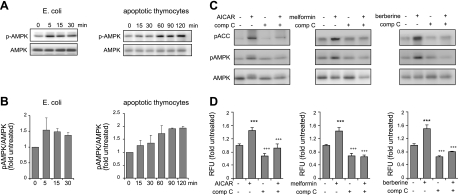
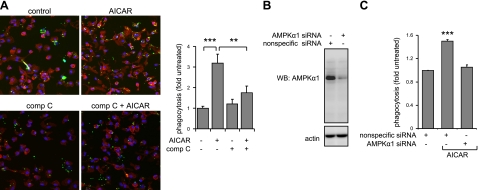
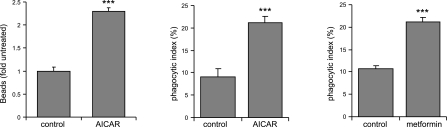
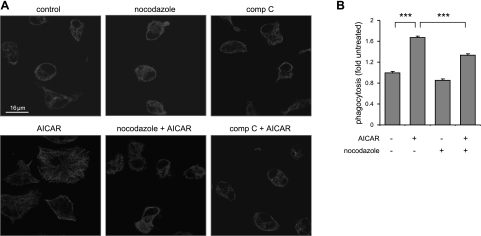
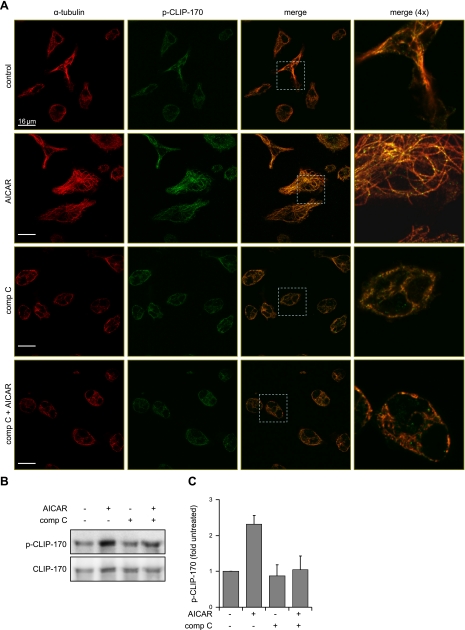
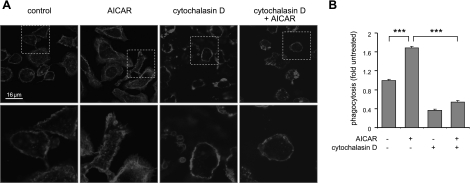
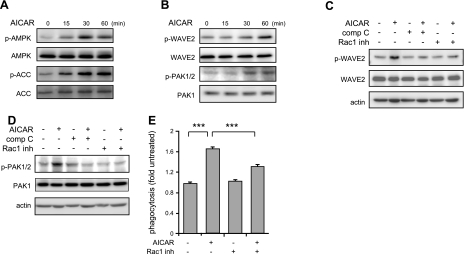
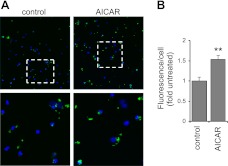
Although AMPK plays well-established roles in the modulation of energy balance, recent studies have shown that AMPK activation has potent anti-inflammatory effects. In the present experiments, we examined the role of AMPK in phagocytosis. We found that ingestion of Escherichia coli or apoptotic cells by macrophages increased AMPK activity. AMPK activation increased the ability of neutrophils or macrophages to ingest bacteria (by 46 ± 7.8 or 85 ± 26%, respectively, compared to control, P<0.05) and the ability of macrophages to ingest apoptotic cells (by 21 ± 1.4%, P<0.05 compared to control). AMPK activation resulted in cytoskeletal reorganization, including enhanced formation of actin and microtubule networks. Activation of PAK1/2 and WAVE2, which are downstream effectors of Rac1, accompanied AMPK activation. AMPK activation also induced phosphorylation of CLIP-170, a protein that participates in microtubule synthesis. The increase in phagocytosis was reversible by the specific AMPK inhibitor compound C, siRNA to AMPKα1, Rac1 inhibitors, or agents that disrupt actin or microtubule networks. In vivo, AMPK activation resulted in enhanced phagocytosis of bacteria in the lungs by 75 ± 5% vs. control (P<0.05). These results demonstrate a novel function for AMPK in enhancing the phagocytic activity of neutrophils and macrophages.
Figures
References
-
- Iseli T. J., Oakhill J. S., Bailey M. F., Wee S., Walter M., van Denderen B. J., Castelli L. A., Katsis F., Witters L. A., Stapleton D., Macaulay S. L., Michell B. J., Kemp B. E. (2008) AMP-activated protein kinase subunit interactions: beta1:gamma1 association requires beta1 Thr-263 and Tyr-267. J. Biol. Chem. 283, 4799–4807 - PubMed
-
- Steinberg G. R., Kemp B. E. (2009) AMPK in health and disease. Physiol. Rev. 89, 1025–1078 - PubMed
-
- Towler M. C., Hardie D. G. (2007) AMP-activated protein kinase in metabolic control and insulin signaling. Circ. Res. 100, 328–341 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
