

CCK stimulation of GLP-1 neurons involves α1-adrenoceptor-mediated increase in glutamatergic synaptic inputs
- PMID: 21885869
- PMCID: PMC3198097
- DOI: 10.2337/db11-0489
CCK stimulation of GLP-1 neurons involves α1-adrenoceptor-mediated increase in glutamatergic synaptic inputs
Abstract
Objective: Glucagon-like peptide 1 (GLP-1) is involved in the central regulation of food intake. It is produced within the brain by preproglucagon (PPG) neurons, which are located primarily within the brain stem. These neurons project widely throughout the brain, including to the appetite centers in the hypothalamus, and are believed to convey signals related to satiety. Previous work demonstrated that they are directly activated by leptin and electrical activity of the afferent vagus. Another satiety hormone, cholecystokinin (CCK), has also been linked to activation of brain stem neurons, suggesting that it might act partially via centrally projecting neurons from the nucleus tractus solitarius (NTS). The aim of this study was to investigate the neuronal circuitry linking CCK to the population of NTS-PPG neurons.
Research design and methods: Transgenic mice expressing yellow fluorescent protein (Venus) under the control of the PPG promoter were used to identify PPG neurons in vitro and to record their electrical and pharmacological profile.
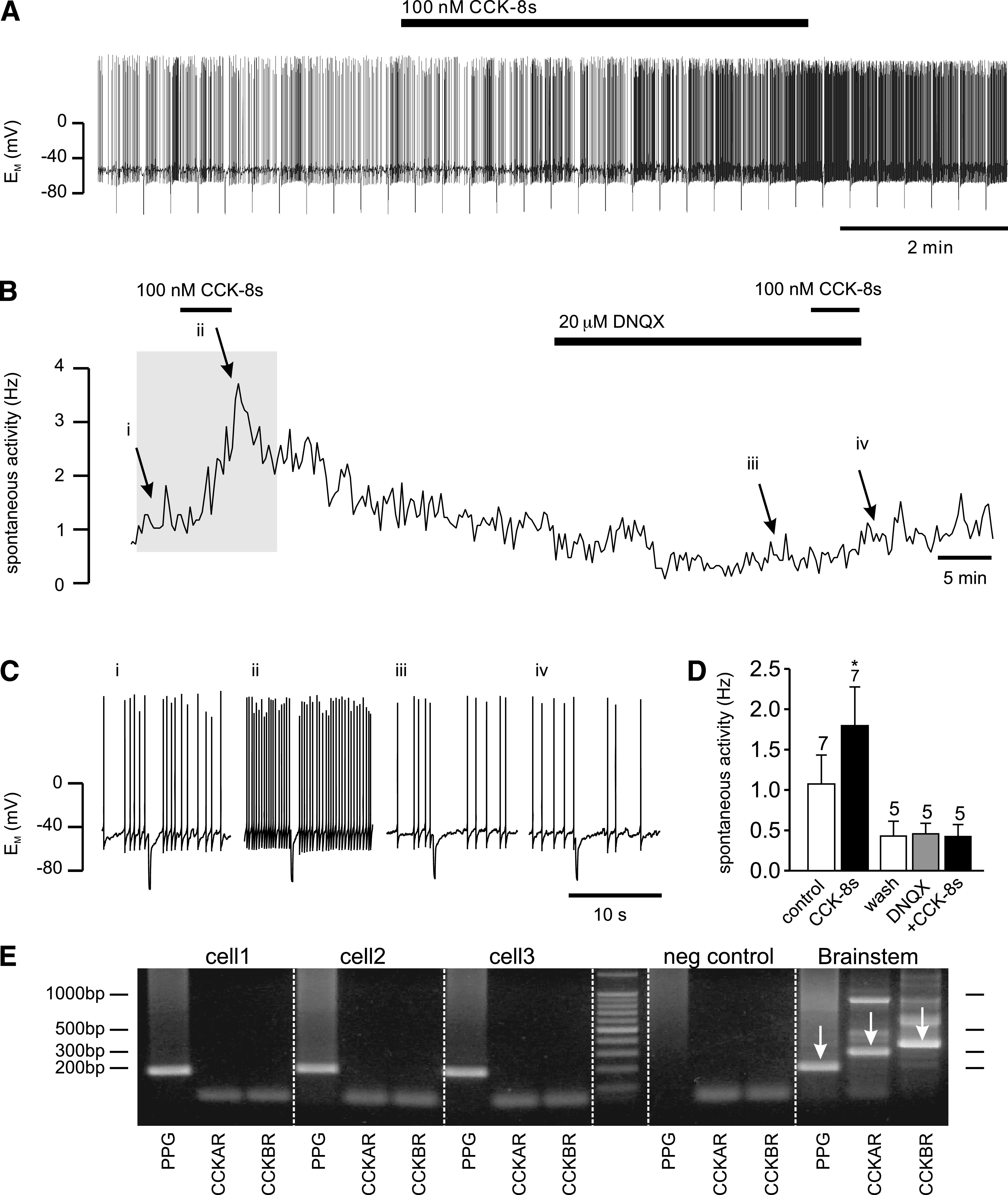
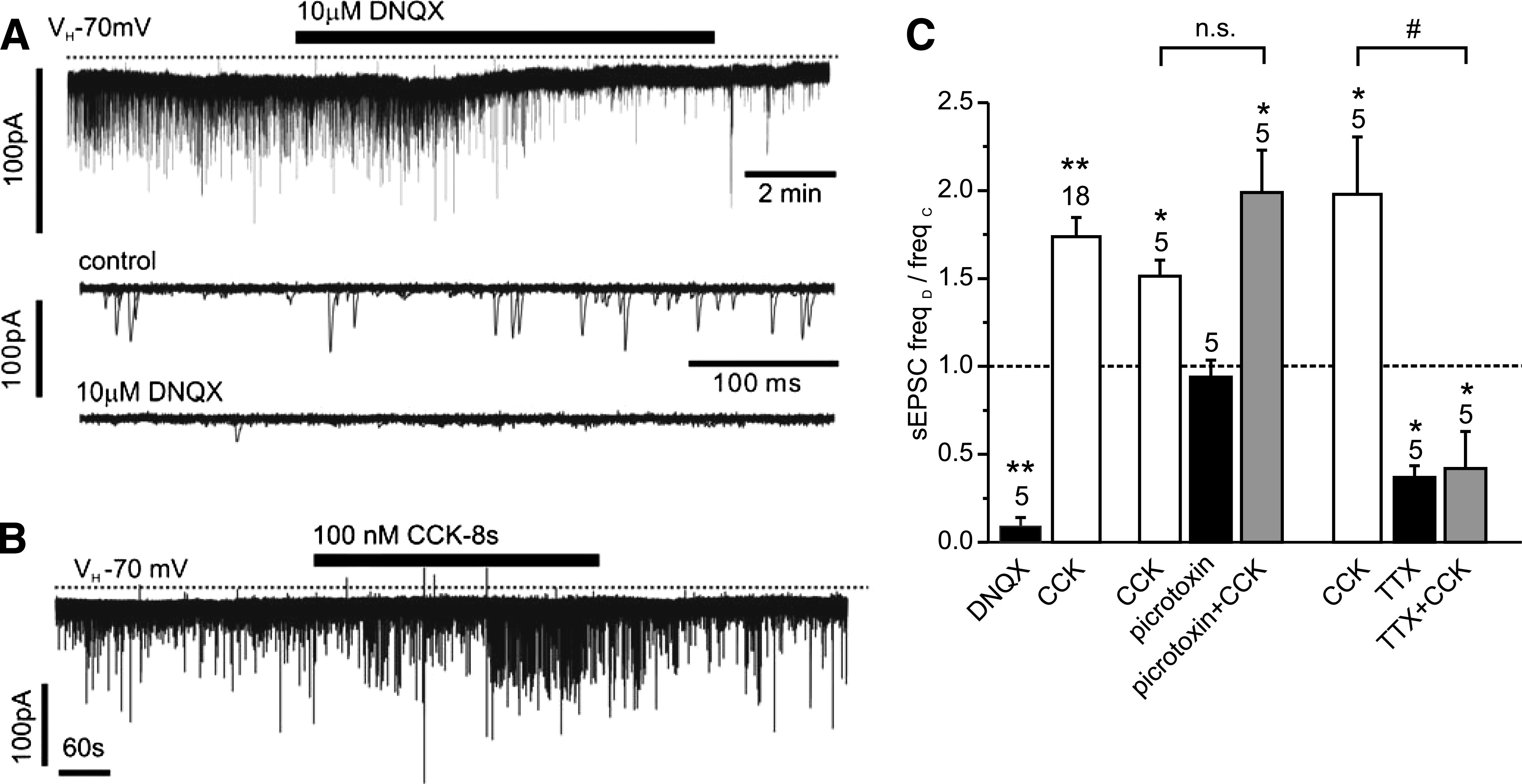
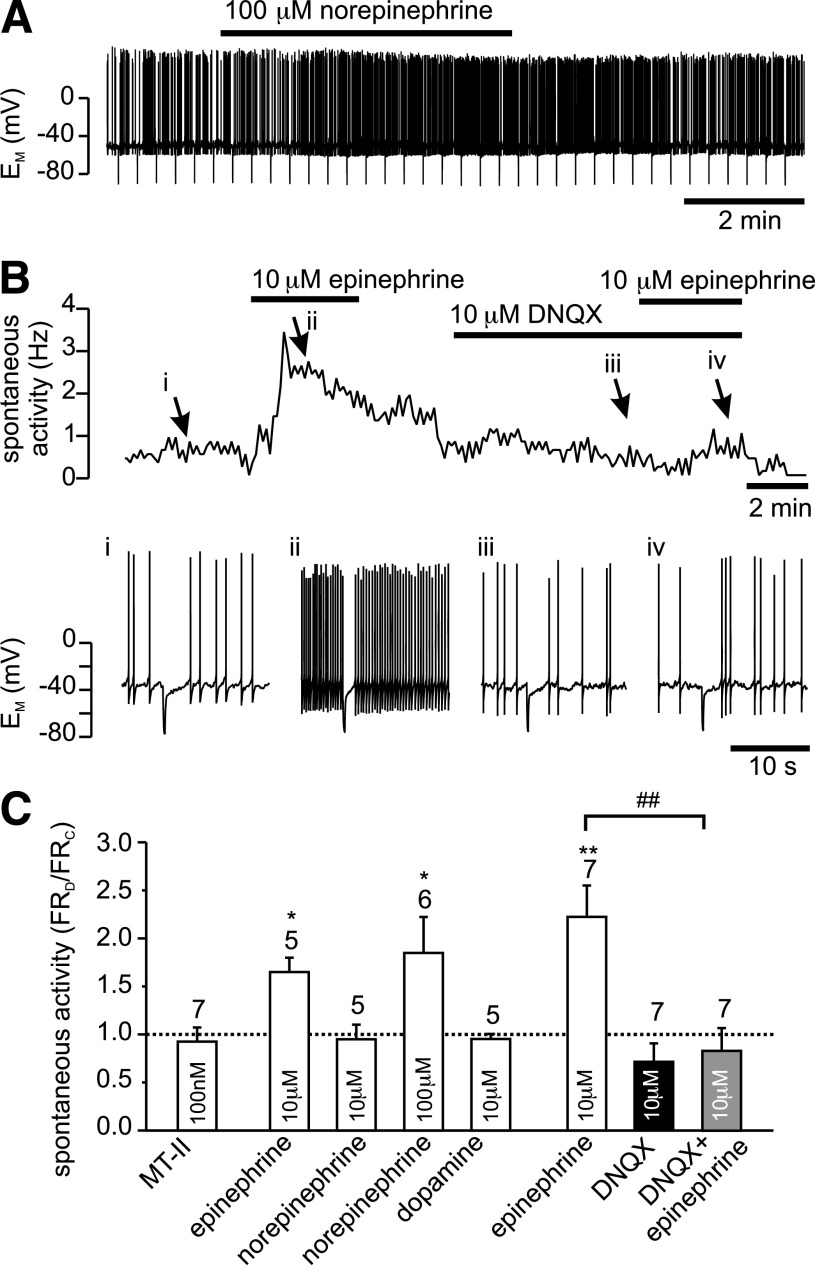
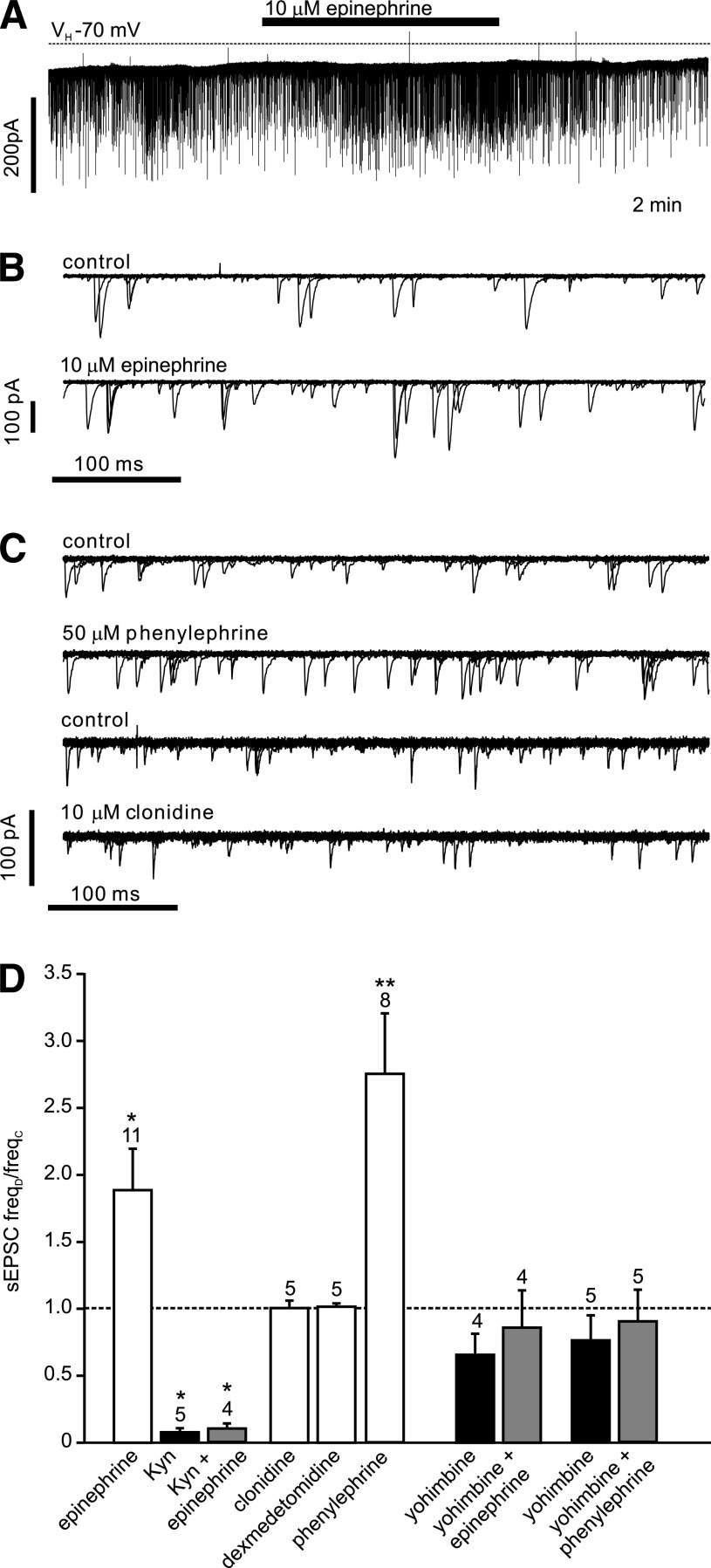
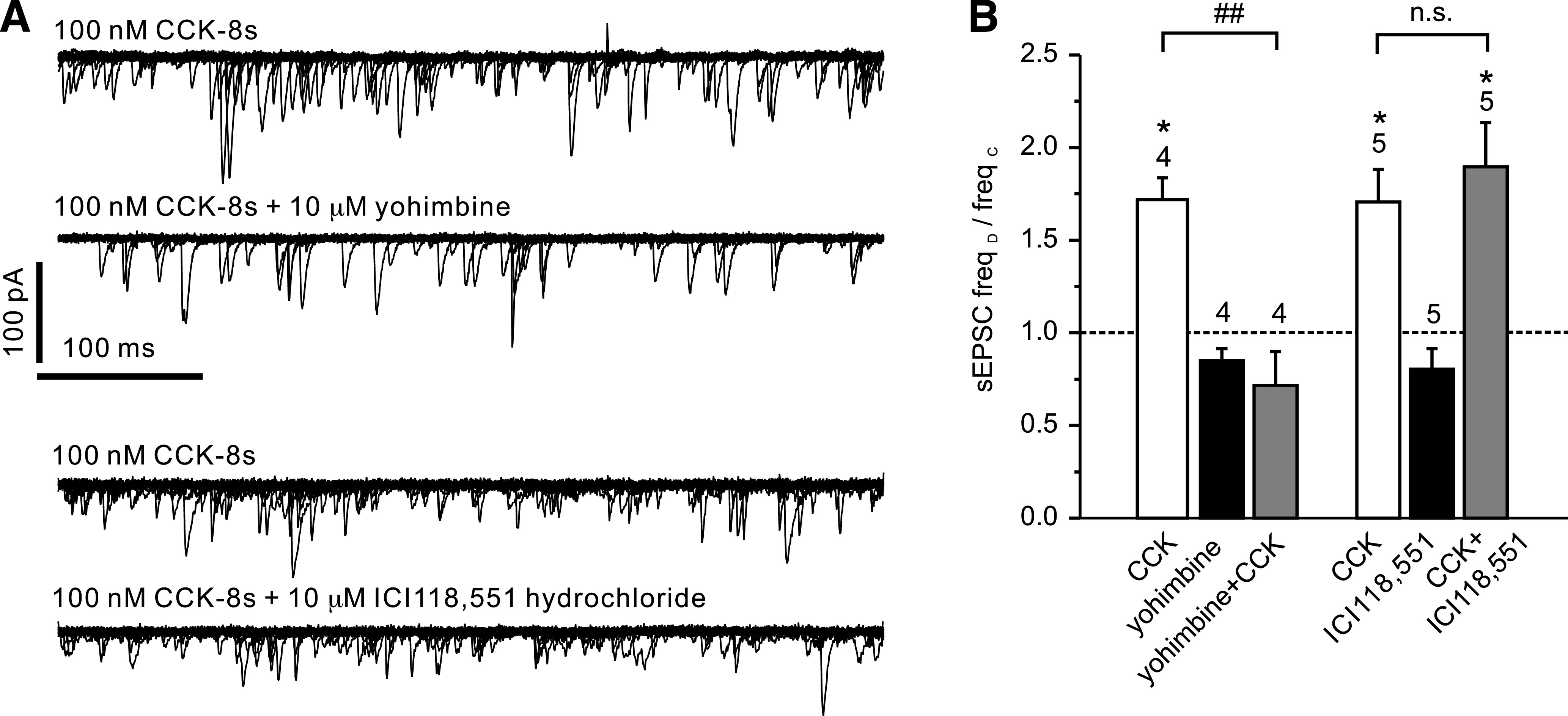
Results: PPG neurons in the NTS were excited by CCK and epinephrine, but not by the melanocortin receptor agonist melanotan II. Both CCK and epinephrine acted to increase glutamatergic transmission to the PPG neurons, and this involved activation of α(1)-adrenergic receptors. Inhibition of adrenergic signaling abolished the excitatory action of CCK.
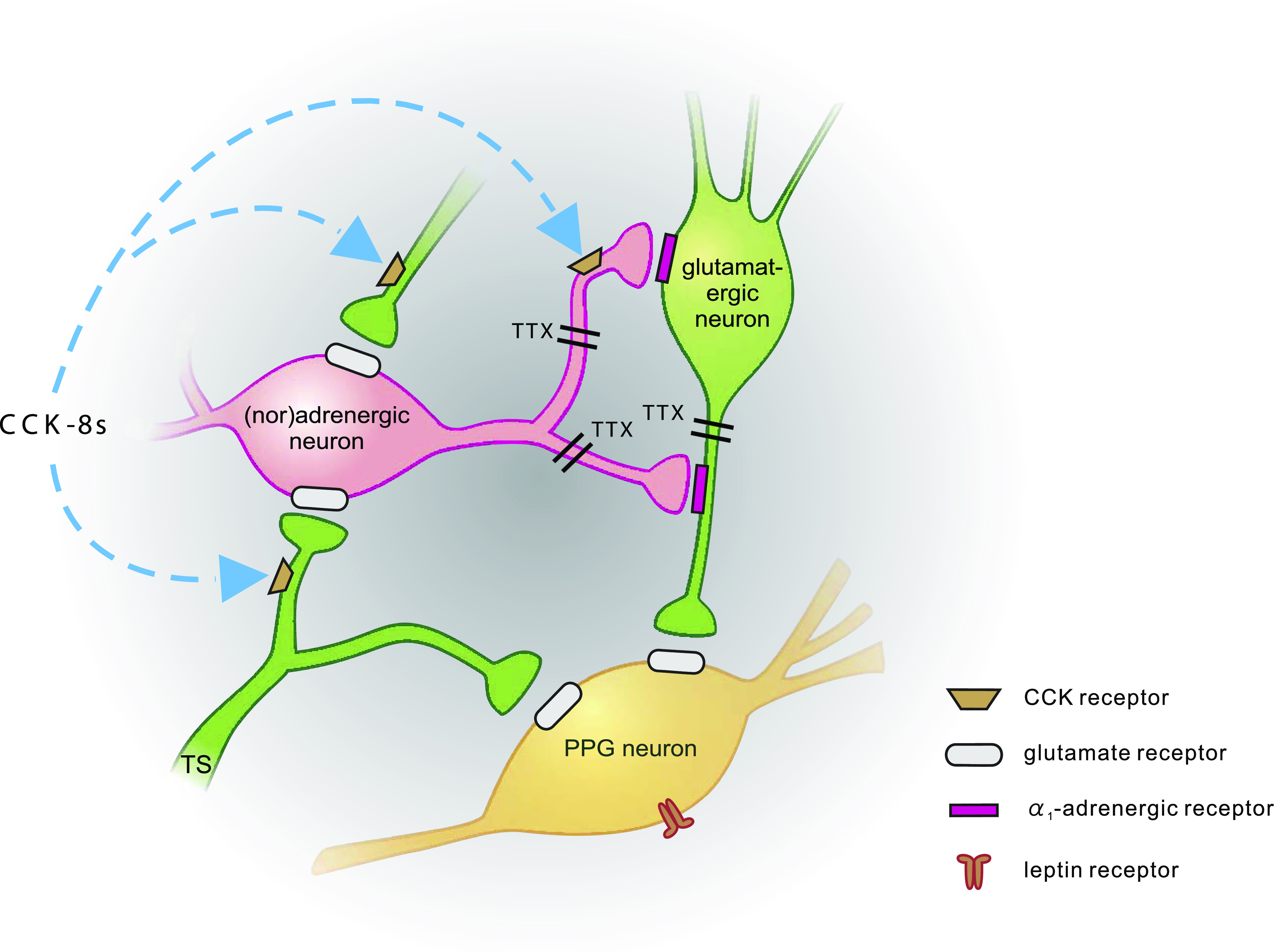
Conclusions: CCK activates NTS-PPG cells by a circuit involving adrenergic and glutamatergic neurons. NTS-PPG neurons integrate a variety of peripheral signals that indicate both long-term energy balance and short-term nutritional and digestional status to produce an output signal to feeding and autonomic circuits.
Figures
References
-
- Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev 2007;87:1409–1439 - PubMed
-
- Trapp S, Hisadome K. Glucagon-like peptide 1 and the brain: central actions-central sources? Auton Neurosci 2011;161:14–19 - PubMed
-
- Merchenthaler I, Lane M, Shughrue P. Distribution of pre-pro-glucagon and glucagon-like peptide-1 receptor messenger RNAs in the rat central nervous system. J Comp Neurol 1999;403:261–280 - PubMed
-
- Jin SL, Han VK, Simmons JG, Towle AC, Lauder JM, Lund PK. Distribution of glucagonlike peptide I (GLP-I), glucagon, and glicentin in the rat brain: an immunocytochemical study. J Comp Neurol 1988;271:519–532 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
