

A dp53/JNK-dependant feedback amplification loop is essential for the apoptotic response to stress in Drosophila
- PMID: 21886179
- PMCID: PMC3278728
- DOI: 10.1038/cdd.2011.113
A dp53/JNK-dependant feedback amplification loop is essential for the apoptotic response to stress in Drosophila
Abstract
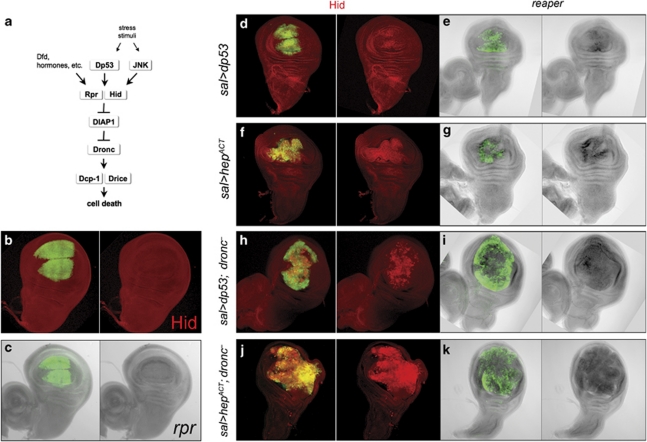
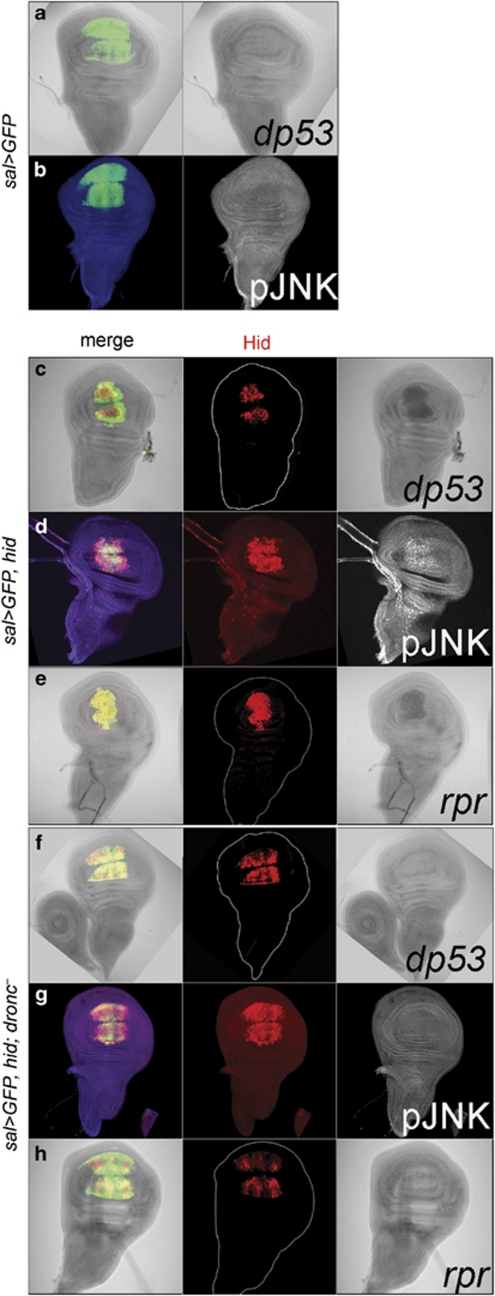
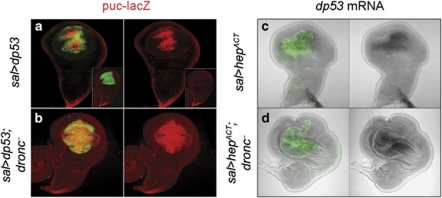
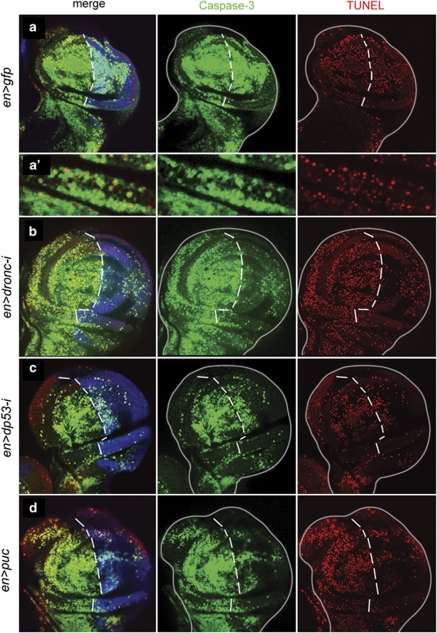
Programmed cell death (apoptosis) is a conserved process aimed to eliminate unwanted cells. The key molecules are a group of proteases called caspases that cleave vital proteins, which leads to the death of cells. In Drosophila, the apoptotic pathway is usually represented as a cascade of events in which an initial stimulus activates one or more of the proapoptotic genes (hid, rpr, grim), which in turn activate caspases. In stress-induced apoptosis, the dp53 (Drosophila p53) gene and the Jun N-terminal kinase (JNK) pathway function upstream in the activation of the proapoptotic genes. Here we demonstrate that dp53 and JNK also function downstream of proapoptotic genes and the initiator caspase Dronc (Drosophila NEDD2-like caspase) and that they establish a feedback loop that amplifies the initial apoptotic stimulus. This loop plays a critical role in the apoptotic response because in its absence there is a dramatic decrease in the amount of cell death after a pulse of the proapoptotic proteins Hid and Rpr. Thus, our results indicate that stress-induced apoptosis in Drosophila is dependant on an amplification loop mediated by dp53 and JNK. Furthermore, they also demonstrate a mechanism of mutual activation of proapoptotic genes.
Figures
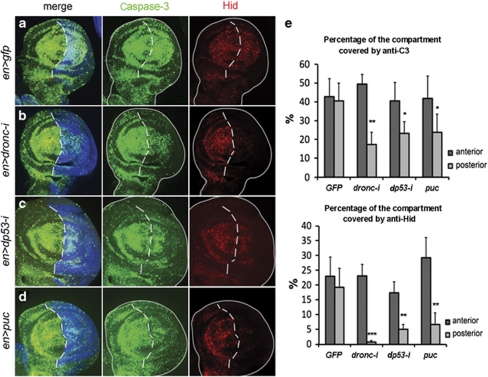
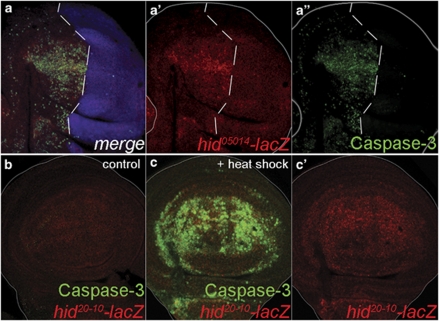
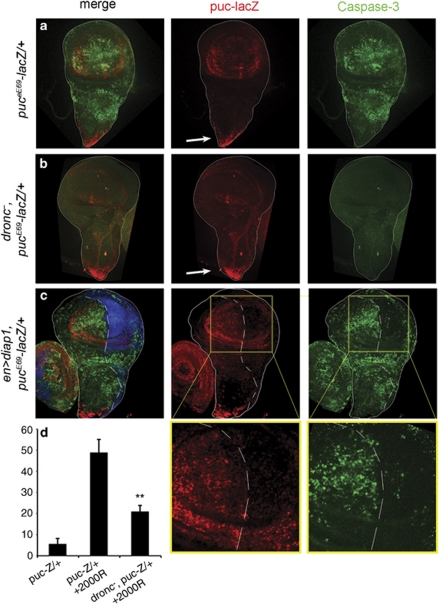
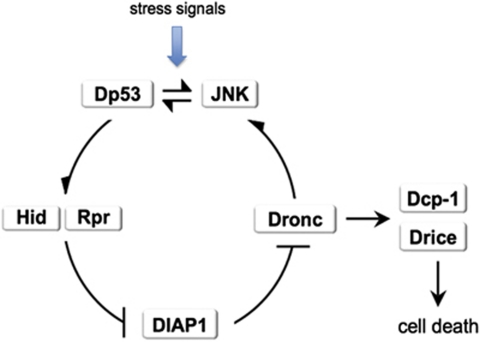
References
-
- Jacobson MD, Weil M, Raff MC. Programmed cell death in animal development. Cell. 1997;88:347–354. - PubMed
-
- Meier P, Finch A, Evan G. Apoptosis in development. Nature. 2000;407:796–801. - PubMed
-
- Thornberry NA, Lazebnik Y. Caspases: enemies within. Science. 1998;281:1312–1316. - PubMed
-
- Reed JC, Doctor KS, Godzik A. The domains of apoptosis: a genomics perspective. Sci STKE. 2004;2004:re9. - PubMed
-
- Shiozaki EN, Shi Y. Caspases, IAPs and Smac/DIABLO: mechanisms from structural biology. Trends Biochem Sci. 2004;29:486–494. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
