

Immunoproteasome-deficiency has no effects on NK cell education, but confers lymphocytes into targets for NK cells in infected wild-type mice
- PMID: 21887316
- PMCID: PMC3161060
- DOI: 10.1371/journal.pone.0023769
Immunoproteasome-deficiency has no effects on NK cell education, but confers lymphocytes into targets for NK cells in infected wild-type mice
Abstract
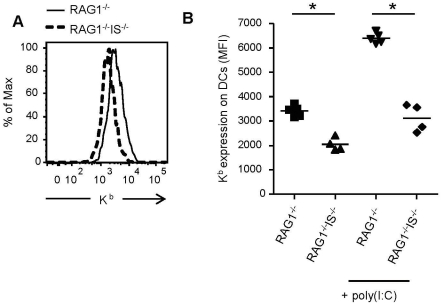
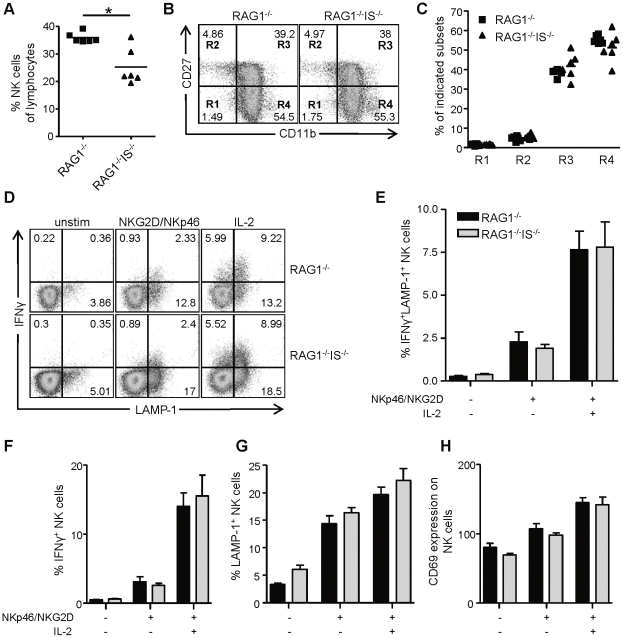
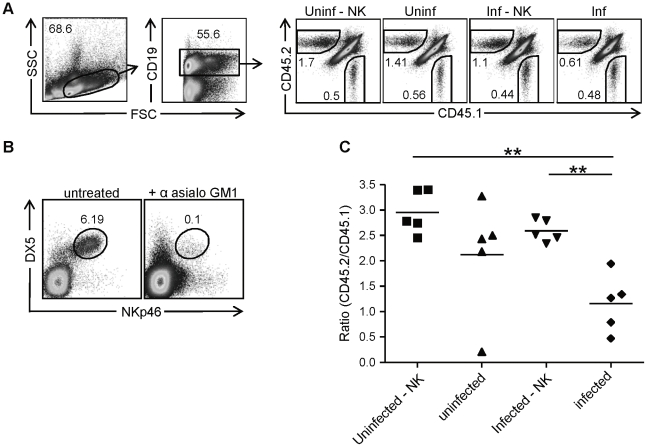
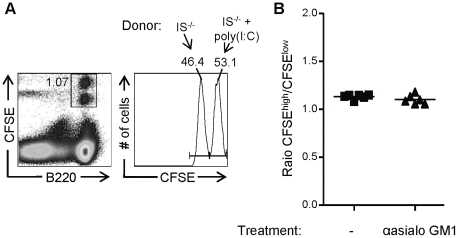
Natural killer (NK) cells are part of the innate immune system and contribute to the eradication of virus infected cells and tumors. NK cells express inhibitory and activating receptors and their decision to kill a target cell is based on the balance of signals received through these receptors. MHC class I molecules are recognized by inhibitory receptors, and their presence during NK cell education influences the responsiveness of peripheral NK cells. We here demonstrate that mice with reduced MHC class I cell surface expression, due to deficiency of immunoproteasomes, have responsive NK cells in the periphery, indicating that the lower MHC class I levels do not alter NK cell education. Following adoptive transfer into wild-type (wt) recipients, immunoproteasome-deficient splenocytes are tolerated in naive but rejected in virus-infected recipients, in an NK cell dependent fashion. These results indicate that the relatively low MHC class I levels are sufficient to protect these cells from rejection by wt NK cells, but that this tolerance is broken in infection, inducing an NK cell-dependent rejection of immunoproteasome-deficient cells.
Conflict of interest statement
Figures
References
-
- Karre K, Ljunggren HG, Piontek G, Kiessling R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature. 1986;319(6055):675–678. - PubMed
-
- Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature. 2005;436(7051):709–713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
