

Myosin IIb activity and phosphorylation status determines dendritic spine and post-synaptic density morphology
- PMID: 21887379
- PMCID: PMC3162601
- DOI: 10.1371/journal.pone.0024149
Myosin IIb activity and phosphorylation status determines dendritic spine and post-synaptic density morphology
Abstract
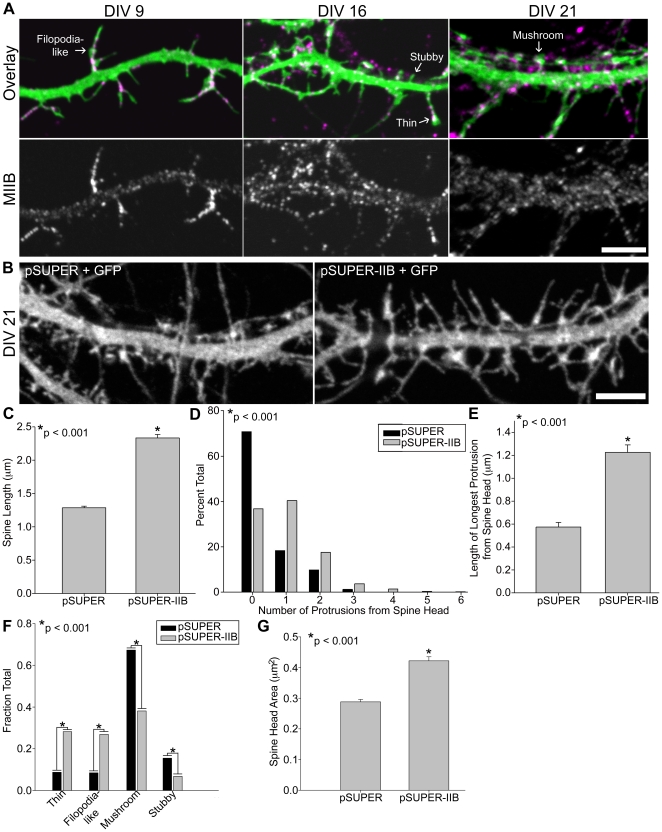
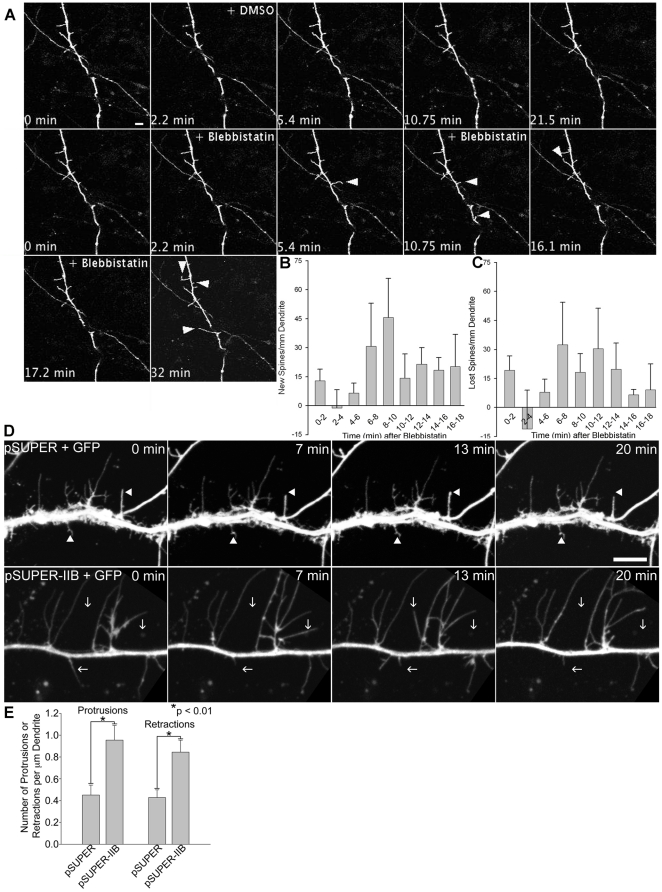
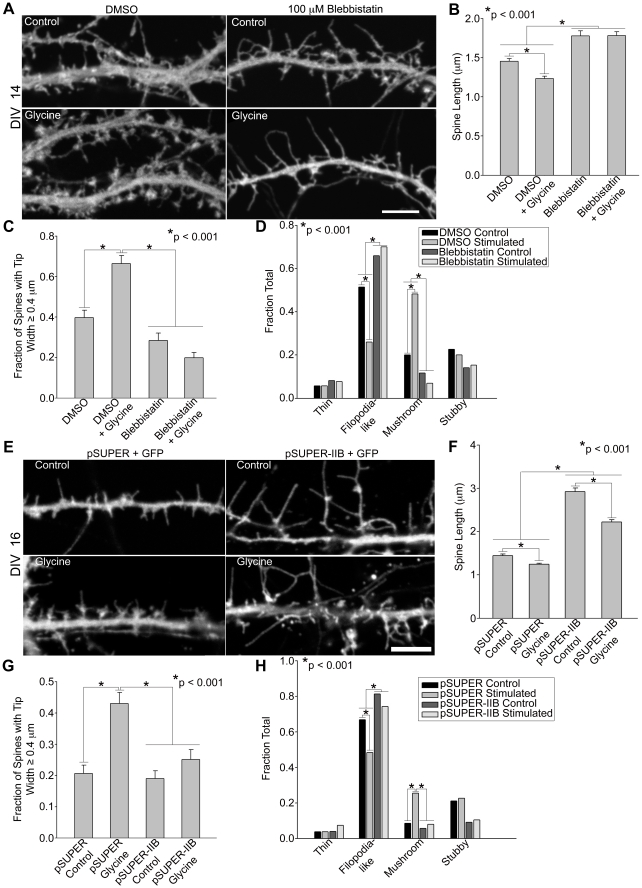
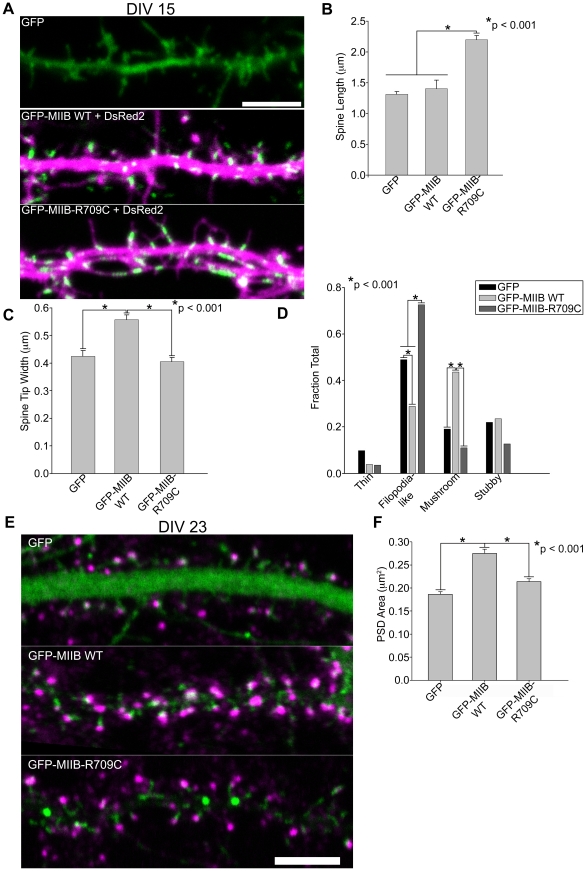
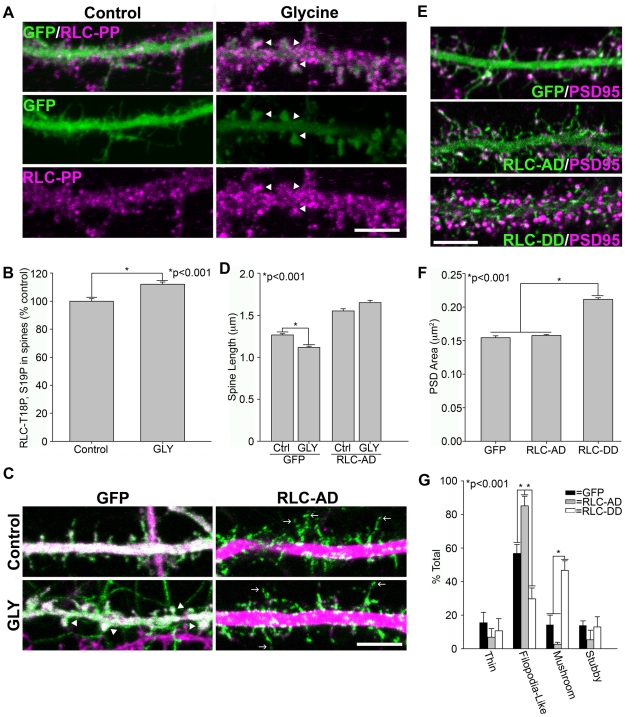
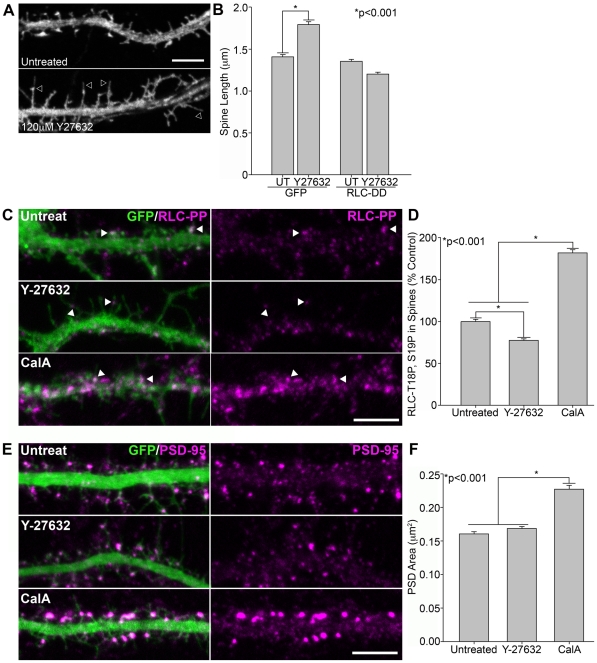
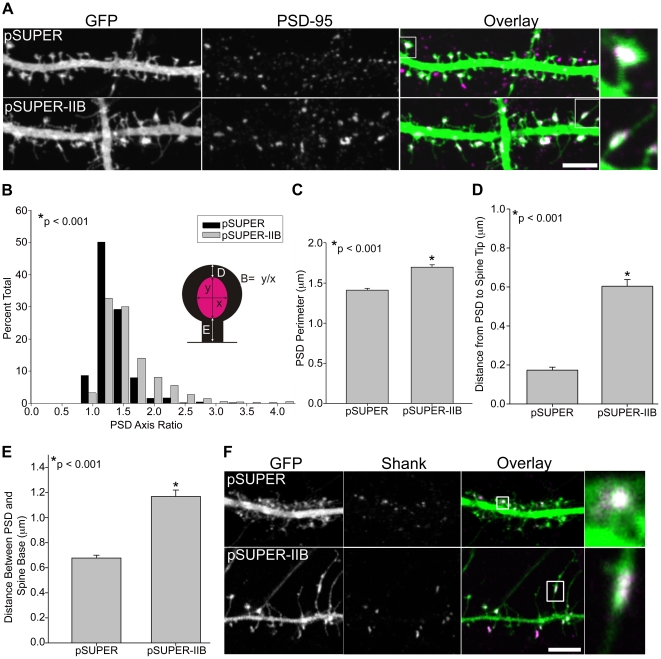
Dendritic spines in hippocampal neurons mature from a filopodia-like precursor into a mushroom-shape with an enlarged post-synaptic density (PSD) and serve as the primary post-synaptic location of the excitatory neurotransmission that underlies learning and memory. Using myosin II regulatory mutants, inhibitors, and knockdowns, we show that non-muscle myosin IIB (MIIB) activity determines where spines form and whether they persist as filopodia-like spine precursors or mature into a mushroom-shape. MIIB also determines PSD size, morphology, and placement in the spine. Local inactivation of MIIB leads to the formation of filopodia-like spine protrusions from the dendritic shaft. However, di-phosphorylation of the regulatory light chain on residues Thr18 and Ser19 by Rho kinase is required for spine maturation. Inhibition of MIIB activity or a mono-phosphomimetic mutant of RLC similarly prevented maturation even in the presence of NMDA receptor activation. Expression of an actin cross-linking, non-contractile mutant, MIIB R709C, showed that maturation into a mushroom-shape requires contractile activity. Loss of MIIB also leads to an elongated PSD morphology that is no longer restricted to the spine tip; whereas increased MIIB activity, specifically through RLC-T18, S19 di-phosphorylation, increases PSD area. These observations support a model whereby myosin II inactivation forms filopodia-like protrusions that only mature once NMDA receptor activation increases RLC di-phosphorylation to stimulate MIIB contractility, resulting in mushroom-shaped spines with an enlarged PSD.
Conflict of interest statement
Figures
References
-
- Sekino Y, Kojima N, Shirao T. Role of actin cytoskeleton in dendritic spine morphogenesis. Neurochem Int. 2007;51:92–104. - PubMed
-
- Yuste R, Bonhoeffer T. Genesis of dendritic spines: insights from ultrastructural and imaging studies. Nat Rev Neurosci. 2004;5:24–34. - PubMed
-
- Peng J, Kim MJ, Cheng D, Duong DM, Gygi SP, et al. Semiquantitative proteomic analysis of rat forebrain postsynaptic density fractions by mass spectrometry. J Biol Chem. 2004;279:21003–21011. - PubMed
-
- Sheng M, Hoogenraad CC. The Postsynaptic Architecture of Excitatory Synapses: A More Quantitative View. Annual Review of Biochemistry. 2007;76:823–847. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
