

Microsecond unfolding kinetics of sheep prion protein reveals an intermediate that correlates with susceptibility to classical scrapie
- PMID: 21889460
- PMCID: PMC3164147
- DOI: 10.1016/j.bpj.2011.07.024
Microsecond unfolding kinetics of sheep prion protein reveals an intermediate that correlates with susceptibility to classical scrapie
Abstract
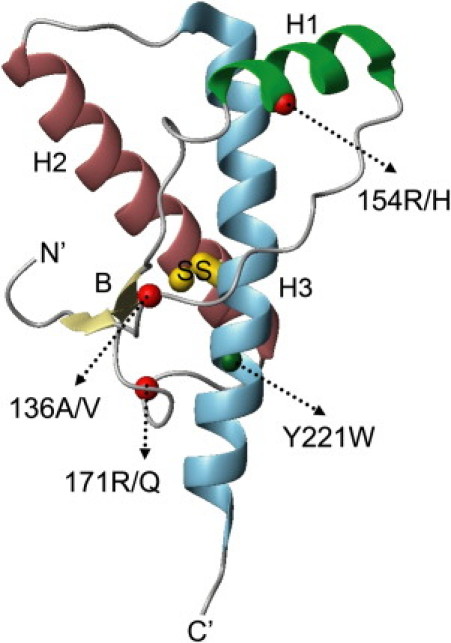
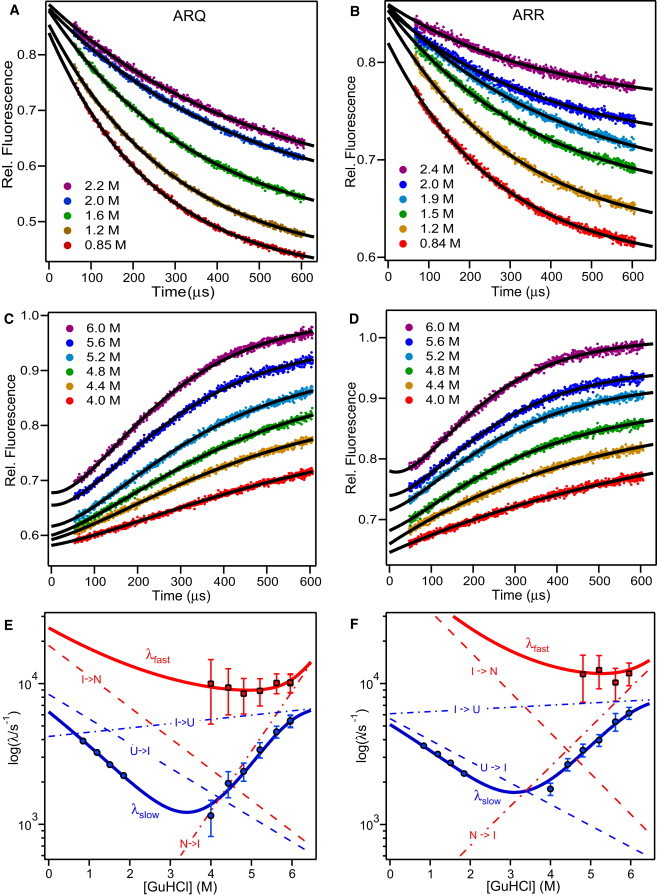
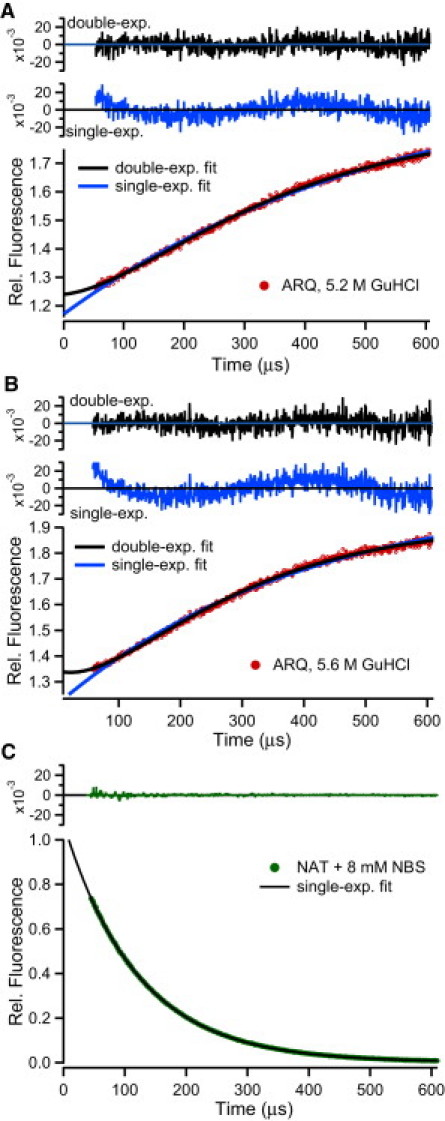
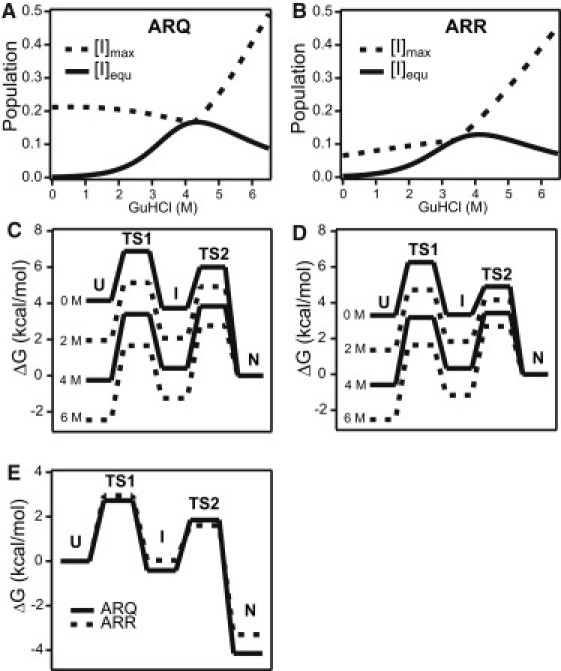
The microsecond folding and unfolding kinetics of ovine prion proteins (ovPrP) were measured under various solution conditions. A fragment comprising residues 94-233 of the full-length ovPrP was studied for four variants with differing susceptibilities to classical scrapie in sheep. The observed biexponential unfolding kinetics of ovPrP provides evidence for an intermediate species. However, in contrast to previous results for human PrP, there is no evidence for an intermediate under refolding conditions. Global analysis of the kinetic data, based on a sequential three-state mechanism, quantitatively accounts for all folding and unfolding data as a function of denaturant concentration. The simulations predict that an intermediate accumulates under both folding and unfolding conditions, but is observable only in unfolding experiments because the intermediate is optically indistinguishable from the native state. The relative population of intermediates in two ovPrP variants, both transiently and under destabilizing equilibrium conditions, correlates with their propensities for classical scrapie. The variant susceptible to classical scrapie has a larger population of the intermediate state than the resistant variant. Thus, the susceptible variant should be favored to undergo the PrP(C) to PrP(Sc) conversion and oligomerization.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Prusiner S.B. Novel proteinaceous infectious particles cause scrapie. Science. 1982;216:136–144. - PubMed
-
- Griffith J.S. Self-replication and scrapie. Nature. 1967;215:1043–1044. - PubMed
-
- Aguzzi A., Calella A.M. Prions: protein aggregation and infectious diseases. Physiol. Rev. 2009;89:1105–1152. - PubMed
-
- Brown D.R., Qin K., Kretzschmar H. The cellular prion protein binds copper in vivo. Nature. 1997;390:684–687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
