

STED nanoscopy of actin dynamics in synapses deep inside living brain slices
- PMID: 21889466
- PMCID: PMC3164186
- DOI: 10.1016/j.bpj.2011.07.027
STED nanoscopy of actin dynamics in synapses deep inside living brain slices
Abstract
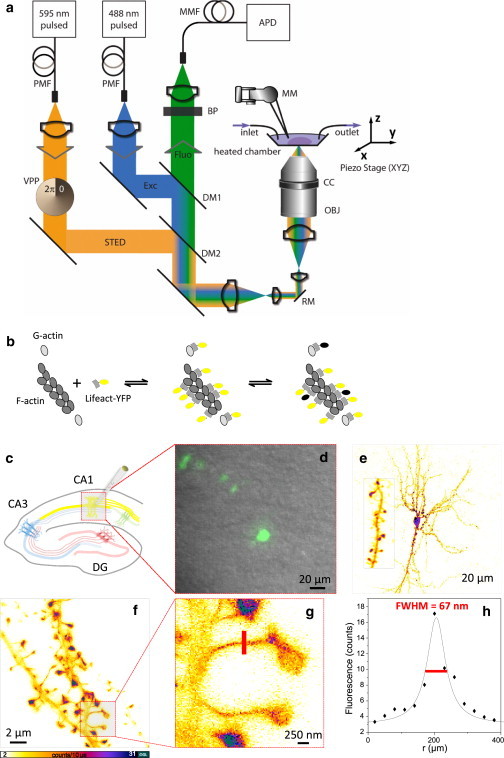
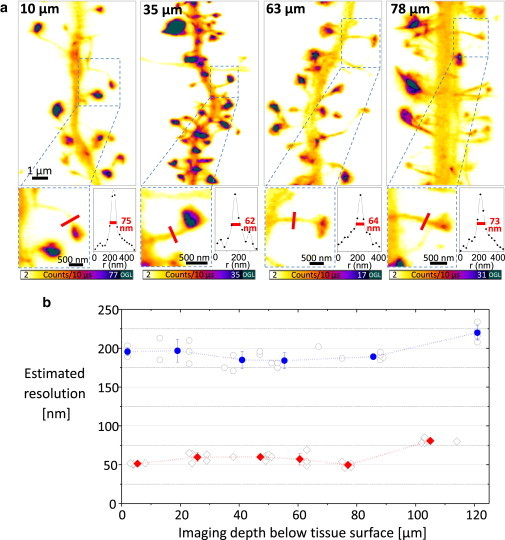
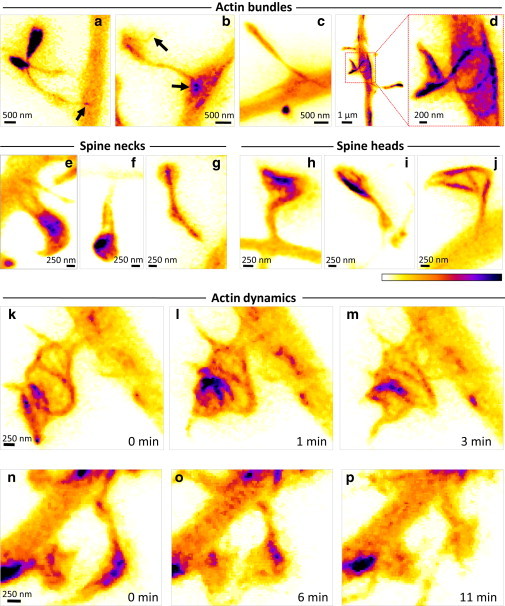
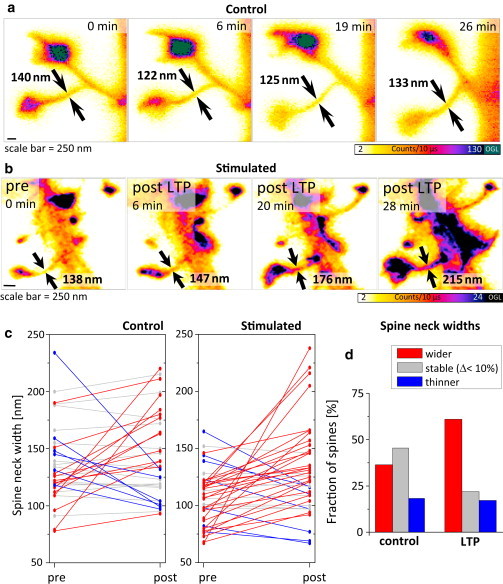
It is difficult to investigate the mechanisms that mediate long-term changes in synapse function because synapses are small and deeply embedded inside brain tissue. Although recent fluorescence nanoscopy techniques afford improved resolution, they have so far been restricted to dissociated cells or tissue surfaces. However, to study synapses under realistic conditions, one must image several cell layers deep inside more-intact, three-dimensional preparations that exhibit strong light scattering, such as brain slices or brains in vivo. Using aberration-reducing optics, we demonstrate that it is possible to achieve stimulated emission depletion superresolution imaging deep inside scattering biological tissue. To illustrate the power of this novel (to our knowledge) approach, we resolved distinct distributions of actin inside dendrites and spines with a resolution of 60-80 nm in living organotypic brain slices at depths up to 120 μm. In addition, time-lapse stimulated emission depletion imaging revealed changes in actin-based structures inside spines and spine necks, and showed that these dynamics can be modulated by neuronal activity. Our approach greatly facilitates investigations of actin dynamics at the nanoscale within functionally intact brain tissue.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Halpain S. Actin and the agile spine: how and why do dendritic spines dance? Trends Neurosci. 2000;23:141–146. - PubMed
-
- Renner M., Specht C.G., Triller A. Molecular dynamics of postsynaptic receptors and scaffold proteins. Curr. Opin. Neurobiol. 2008;18:532–540. - PubMed
-
- Sheng M., Hoogenraad C.C. The postsynaptic architecture of excitatory synapses: a more quantitative view. Annu. Rev. Biochem. 2007;76:823–847. - PubMed
-
- Svoboda K., Yasuda R. Principles of two-photon excitation microscopy and its applications to neuroscience. Neuron. 2006;50:823–839. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
