

AKAP220 protein organizes signaling elements that impact cell migration
- PMID: 21890631
- PMCID: PMC3234751
- DOI: 10.1074/jbc.M111.277756
AKAP220 protein organizes signaling elements that impact cell migration
Abstract
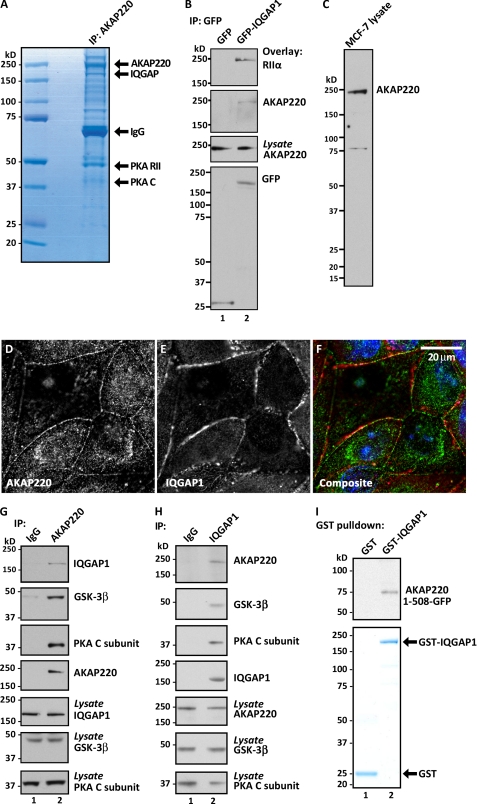
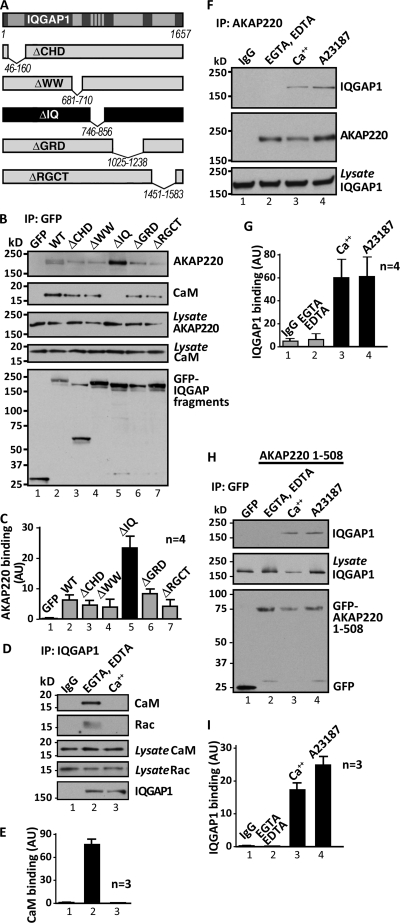
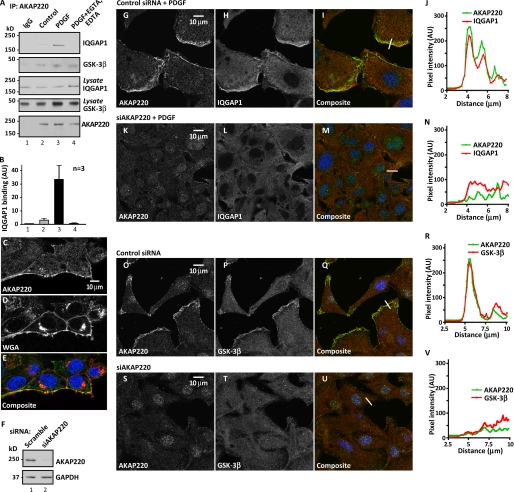
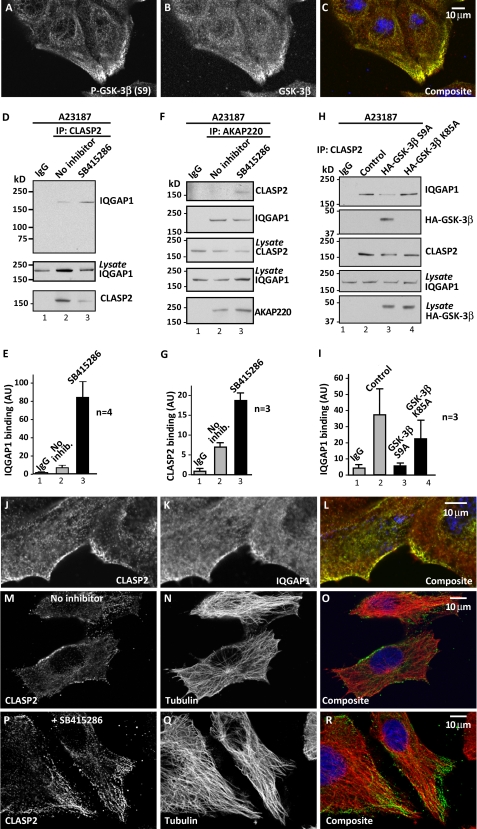
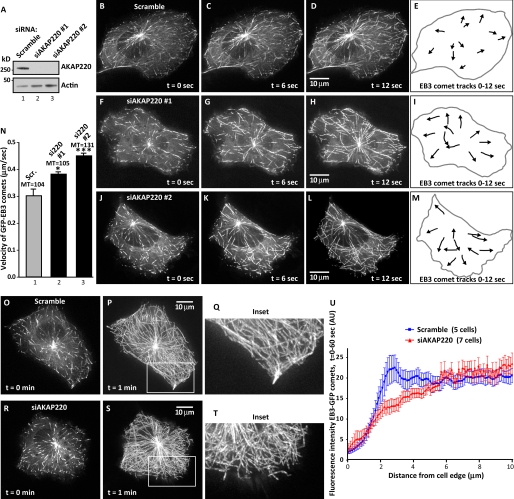
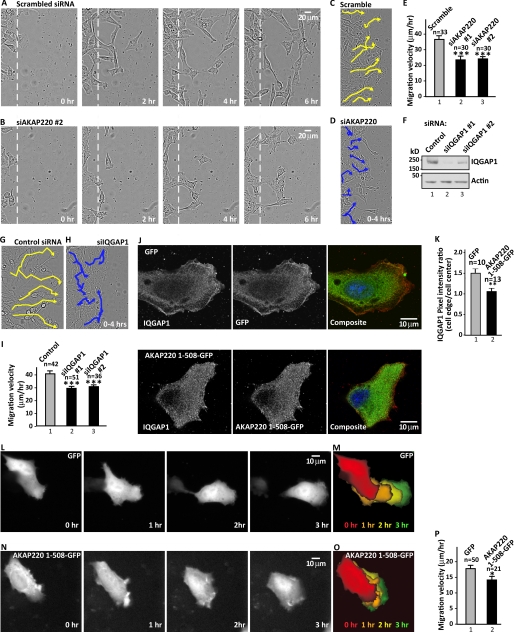
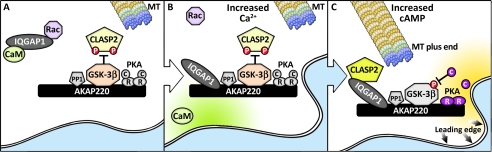
Cell movement requires the coordinated reception, integration, and processing of intracellular signals. We have discovered that the protein kinase A anchoring protein AKAP220 interacts with the cytoskeletal scaffolding protein IQGAP1 to influence cell motility. AKAP220/IQGAP1 networks receive and integrate calcium and cAMP second messenger signals and position signaling enzymes near their intended substrates at leading edges of migrating cells. IQGAP1 supports calcium/calmodulin-dependent association of factors that modulate microtubule dynamics. AKAP220 suppresses GSK-3β and positions this kinase to allow recruitment of the plus-end microtubule tracking protein CLASP2. Gene silencing of AKAP220 alters the rate of microtubule polymerization and the lateral tracking of growing microtubules and retards cell migration in metastatic human cancer cells. This reveals an unappreciated role for this anchored kinase/microtubule effector protein network in the propagation of cell motility.
Figures
References
-
- Pidoux G., Taskén K. (2010) J. Mol. Endocrinol. 44, 271–284 - PubMed
-
- Carr D. W., Stofko-Hahn R. E., Fraser I. D., Bishop S. M., Acott T. S., Brennan R. G., Scott J. D. (1991) J. Biol. Chem. 266, 14188–14192 - PubMed
-
- Gold M. G., Lygren B., Dokurno P., Hoshi N., McConnachie G., Taskén K., Carlson C. R., Scott J. D., Barford D. (2006) Mol. Cell 24, 383–395 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
