

Fitness landscapes: an alternative theory for the dominance of mutation
- PMID: 21890744
- PMCID: PMC3213354
- DOI: 10.1534/genetics.111.132944
Fitness landscapes: an alternative theory for the dominance of mutation
Abstract
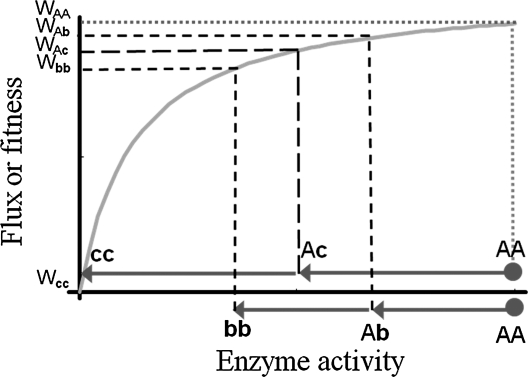
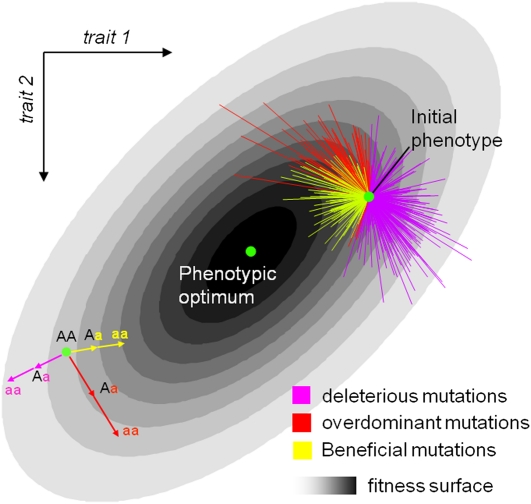
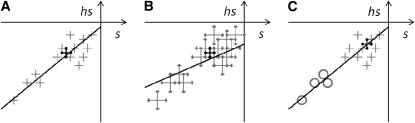
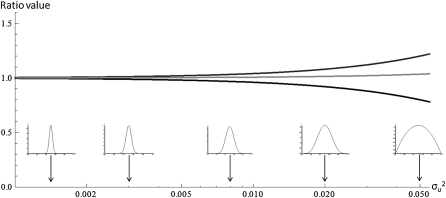
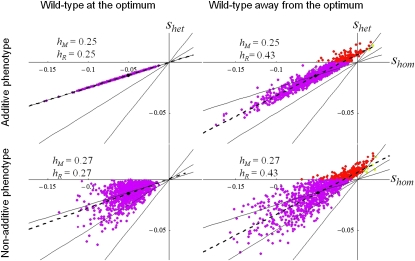
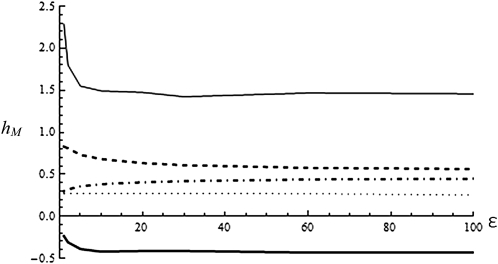
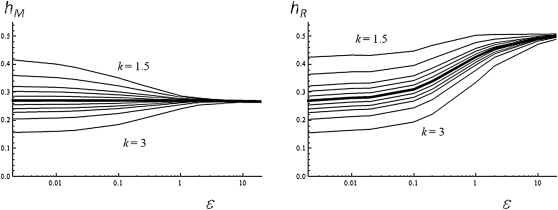
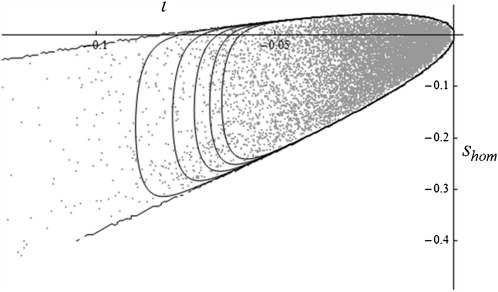
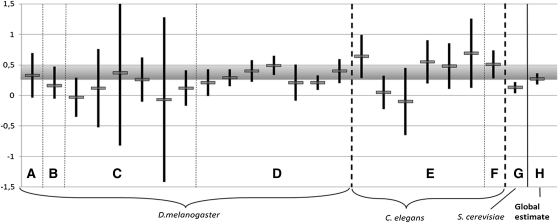
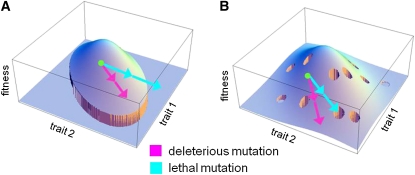
Deleterious mutations tend to be recessive. Several theories, notably those of Fisher (based on selection) and Wright (based on metabolism), have been put forward to explain this pattern. Despite a long-lasting debate, the matter remains unresolved. This debate has focused on the average dominance of mutations. However, we also know very little about the distribution of dominance coefficients among mutations, and about its variation across environments. In this article we present a new approach to predicting this distribution. Our approach is based on a phenotypic fitness landscape model. First, we show that under a very broad range of conditions (and environments), the average dominance of mutation of small effects should be approximately one-quarter as long as adaptation of organisms to their environment can be well described by stabilizing selection on an arbitrary set of phenotypic traits. Second, the theory allows predicting the whole distribution of dominance coefficients among mutants. Because it provides quantitative rather than qualitative predictions, this theory can be directly compared to data. We found that its prediction on mean dominance (average dominance close to 0.25) agreed well with the data, based on a meta-analysis of dominance data for mildly deleterious mutations. However, a simple landscape model does not account for the dominance of mutations of large effects and we provide possible extension of the theory for this class of mutations. Because dominance is a central parameter for evolutionary theory, and because these predictions are quantitative, they set the stage for a wide range of applications and further empirical tests.
Figures
References
-
- Agrawal A. F., 2009. Spatial heterogeneity and the evolution of sex in diploids. Am. Nat. 174: S54–S70 - PubMed
-
- Barton N. H., 2001. The role of hybridization in evolution. Mol. Ecol. 10: 551–568 - PubMed
-
- Bataillon T., Kirkpatrick M., 2000. Inbreeding depression due to mildly deleterious mutations in finite populations: size does matter. Genet. Res. 75: 75–81 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
