

Activity, specificity and structure of I-Bth0305I: a representative of a new homing endonuclease family
- PMID: 21890897
- PMCID: PMC3239194
- DOI: 10.1093/nar/gkr669
Activity, specificity and structure of I-Bth0305I: a representative of a new homing endonuclease family
Abstract
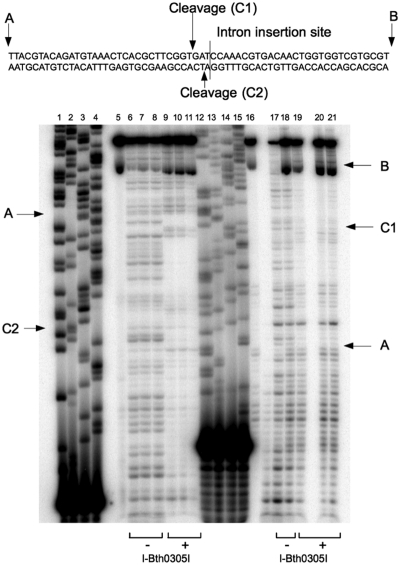
Novel family of putative homing endonuclease genes was recently discovered during analyses of metagenomic and genomic sequence data. One such protein is encoded within a group I intron that resides in the recA gene of the Bacillus thuringiensis 03058-36 bacteriophage. Named I-Bth0305I, the endonuclease cleaves a DNA target in the uninterrupted recA gene at a position immediately adjacent to the intron insertion site. The enzyme displays a multidomain, homodimeric architecture and footprints a DNA region of ~60 bp. Its highest specificity corresponds to a 14-bp pseudopalindromic sequence that is directly centered across the DNA cleavage site. Unlike many homing endonucleases, the specificity profile of the enzyme is evenly distributed across much of its target site, such that few single base pair substitutions cause a significant decrease in cleavage activity. A crystal structure of its C-terminal domain confirms a nuclease fold that is homologous to very short patch repair (Vsr) endonucleases. The domain architecture and DNA recognition profile displayed by I-Bth0305I, which is the prototype of a homing lineage that we term the 'EDxHD' family, are distinct from previously characterized homing endonucleases.
© The Author(s) 2011. Published by Oxford University Press.
Figures







References
-
- Chevalier B, Monnat RJ, Jr, Stoddard BL. In: Homing Endonucleases and Inteins. Belfort M, Wood D, Derbyshire V, Stoddard B, editors. Vol. 16. Berlin: Springer Verlag; 2005. pp. 34–47.
-
- Flick KE, Jurica MS, Monnat RJ, Jr, Stoddard BL. DNA binding and cleavage by the nuclear intron-encoded homing endonuclease I-PpoI. Nature. 1998;394:96–101. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
