

Influence of ground-state structure and Mg2+ binding on folding kinetics of the guanine-sensing riboswitch aptamer domain
- PMID: 21890900
- PMCID: PMC3239184
- DOI: 10.1093/nar/gkr664
Influence of ground-state structure and Mg2+ binding on folding kinetics of the guanine-sensing riboswitch aptamer domain
Abstract
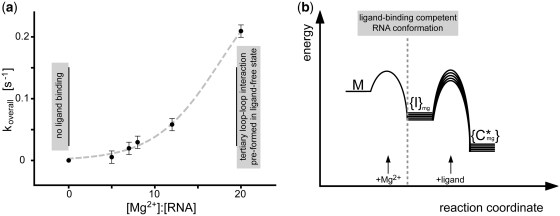
Riboswitch RNAs fold into complex tertiary structures upon binding to their cognate ligand. Ligand recognition is accomplished by key residues in the binding pocket. In addition, it often crucially depends on the stability of peripheral structural elements. The ligand-bound complex of the guanine-sensing riboswitch from Bacillus subtilis, for example, is stabilized by extensive interactions between apical loop regions of the aptamer domain. Previously, we have shown that destabilization of this tertiary loop-loop interaction abrogates ligand binding of the G37A/C61U-mutant aptamer domain (Gsw(loop)) in the absence of Mg(2+). However, if Mg(2+) is available, ligand-binding capability is restored by a population shift of the ground-state RNA ensemble toward RNA conformations with pre-formed loop-loop interactions. Here, we characterize the striking influence of long-range tertiary structure on RNA folding kinetics and on ligand-bound complex structure, both by X-ray crystallography and time-resolved NMR. The X-ray structure of the ligand-bound complex reveals that the global architecture is almost identical to the wild-type aptamer domain. The population of ligand-binding competent conformations in the ground-state ensemble of Gsw(loop) is tunable through variation of the Mg(2+) concentration. We quantitatively describe the influence of distinct Mg(2+) concentrations on ligand-induced folding trajectories both by equilibrium and time-resolved NMR spectroscopy at single-residue resolution.
© The Author(s) 2011. Published by Oxford University Press.
Figures




References
-
- Brion P, Westhof E. Hierarchy and dynamics of RNA folding. Annu. Rev. Biophys. Biomol. Struct. 1997;26:113–137. - PubMed
-
- Draper DE, Grilley D, Soto AM. Ions and RNA folding. Annu. Rev. Biophys. Biomol. Struct. 2005;34:221–243. - PubMed
-
- Thirumalai D, Hyeon C. RNA and protein folding: common themes and variations. Biochemistry. 2005;44:4957–4970. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
