

c-Abl and Arg are activated in human primary melanomas, promote melanoma cell invasion via distinct pathways, and drive metastatic progression
- PMID: 21892207
- PMCID: PMC3235241
- DOI: 10.1038/onc.2011.361
c-Abl and Arg are activated in human primary melanomas, promote melanoma cell invasion via distinct pathways, and drive metastatic progression
Abstract
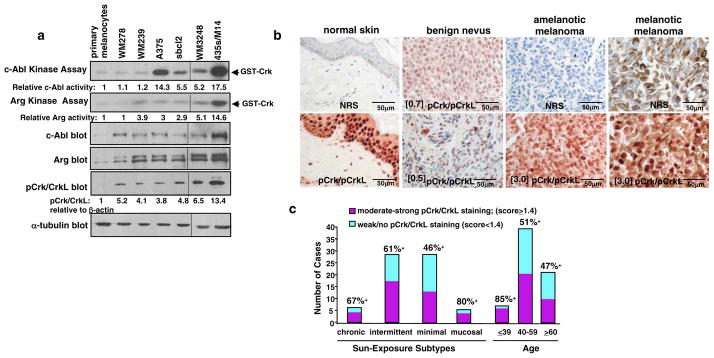
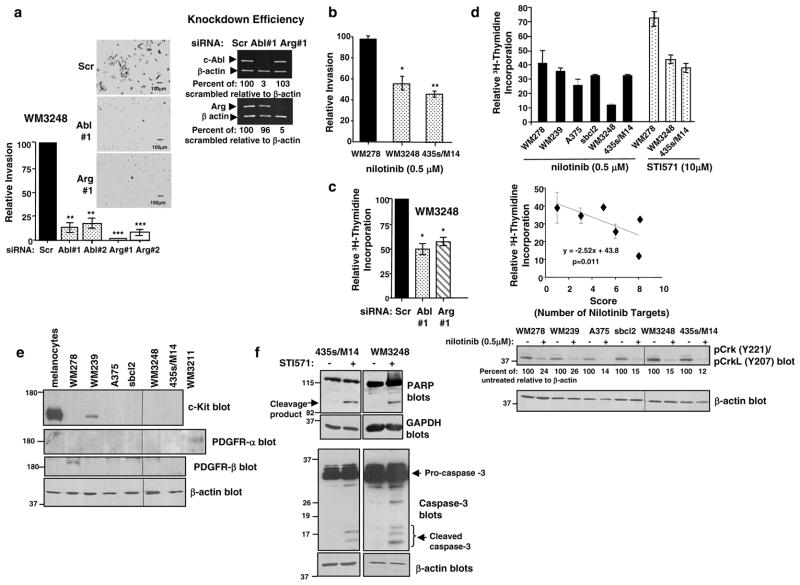
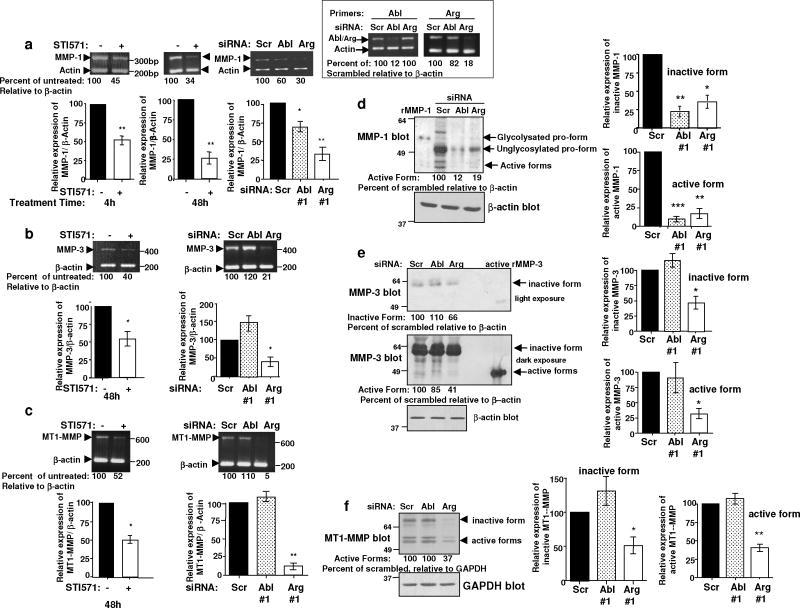
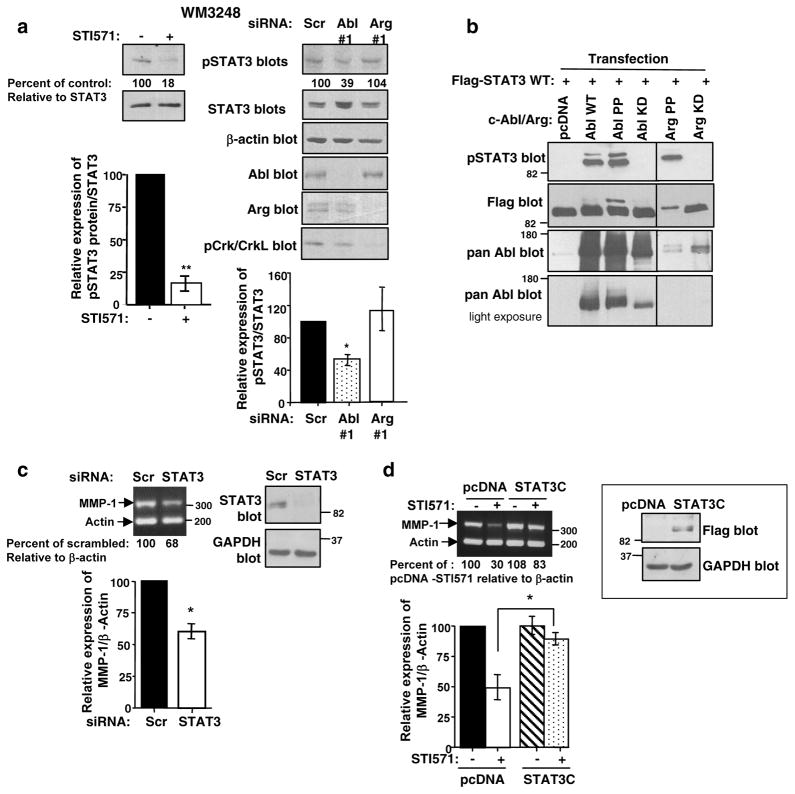
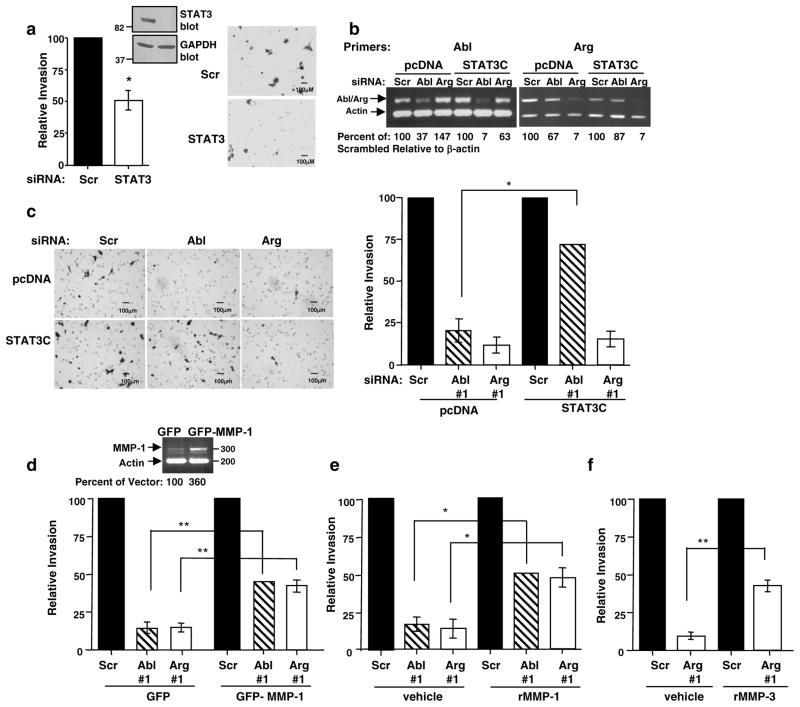
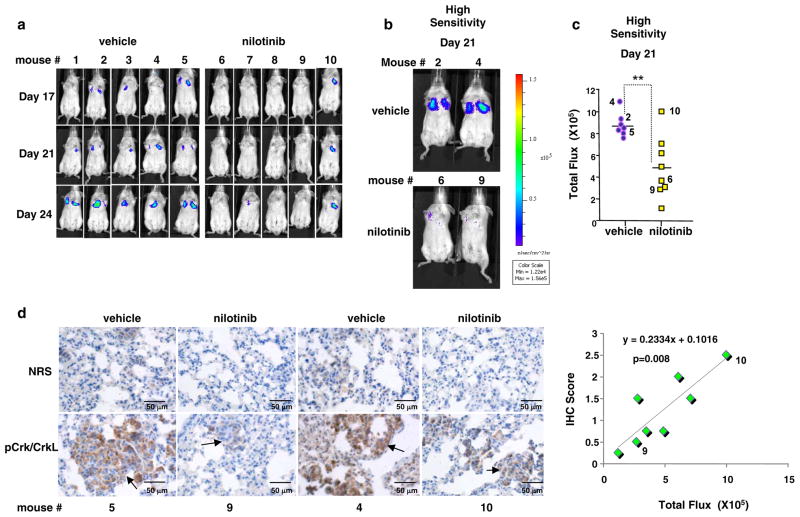
Despite 35 years of clinical trials, there is little improvement in 1-year survival rates for patients with metastatic melanoma, and the disease is essentially untreatable if not cured surgically. The paucity of chemotherapeutic agents that are effective for treating metastatic melanoma indicates a dire need to develop new therapies. Here, we found a previously unrecognized role for c-Abl and Arg in melanoma progression. We demonstrate that the kinase activities of c-Abl and Arg are elevated in primary melanomas (60%), in a subset of benign nevi (33%) and in some human melanoma cell lines. Using siRNA and pharmacological approaches, we show that c-Abl/Arg activation is functionally relevant because it is requiredfor melanoma cell proliferation, survival and invasion. Significantly, we identify the mechanism by which activated c-Abl promotes melanoma invasion by showing that it transcriptionally upregulates matrix metalloproteinase-1 (MMP-1), and using rescue approaches we demonstrate that c-Abl promotes invasion through a STAT3 → MMP-1 pathway. Additionally, we show that c-Abl and Arg are not merely redundant, as active Arg drives invasion in a STAT3-independent manner, and upregulates MMP-3 and MT1-MMP, in addition to MMP-1. Most importantly, c-Abl and Arg not only promote in vitro processes important for melanoma progression, but also promote metastasis in vivo, as inhibition of c-Abl/Arg kinase activity with the c-Abl/Arg inhibitor, nilotinib, dramatically inhibits metastasis in a mouse model. Taken together, these data identify c-Abl and Arg as critical, novel, drug targets in metastatic melanoma, and indicate that nilotinib may be useful in preventing metastasis in patients with melanomas harboring active c-Abl and Arg.
Conflict of interest statement
Figures
References
-
- Bachmeier BE, Albini A, Vene R, Benelli R, Noonan D, Weigert C, et al. Cell density-dependent regulation of matrix metalloproteinase and TIMP expression in differently tumorigenic breast cancer cell lines. Exp Cell Res. 2005;305:83–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
