

The Bam machine: a molecular cooper
- PMID: 21893027
- PMCID: PMC3253334
- DOI: 10.1016/j.bbamem.2011.08.020
The Bam machine: a molecular cooper
Abstract
The bacterial outer membrane (OM) is an exceptional biological structure with a unique composition that contributes significantly to the resiliency of Gram-negative bacteria. Since all OM components are synthesized in the cytosol, the cell must efficiently transport OM-specific lipids and proteins across the cell envelope and stably integrate them into a growing membrane. In this review, we discuss the challenges associated with these processes and detail the elegant solutions that cells have evolved to address the topological problem of OM biogenesis. Special attention will be paid to the Bam machine, a highly conserved multiprotein complex that facilitates OM β-barrel folding. This article is part of a Special Issue entitled: Protein Folding in Membranes.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
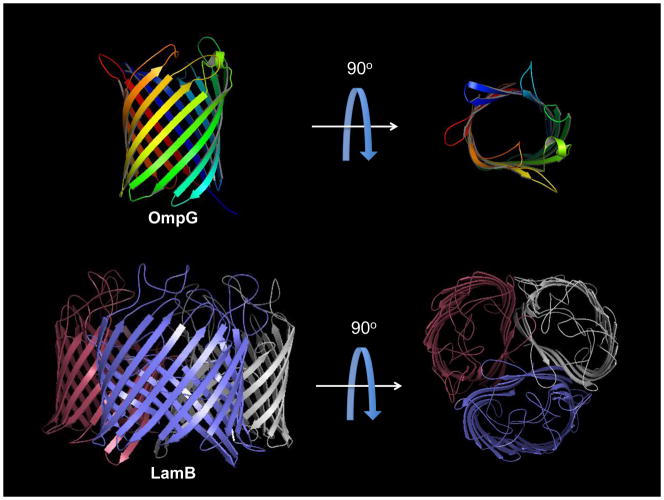
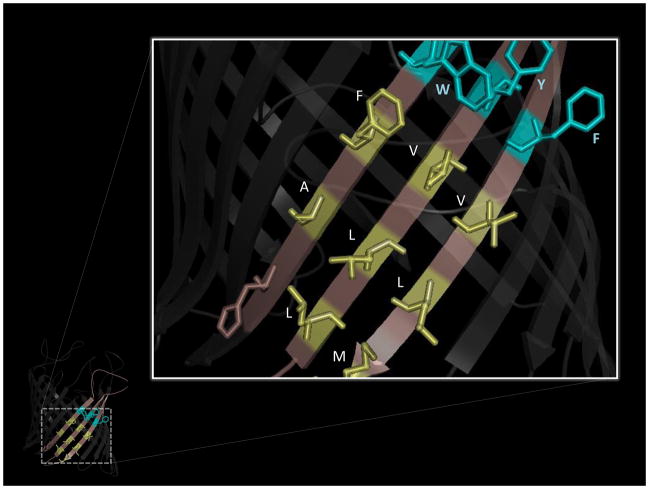
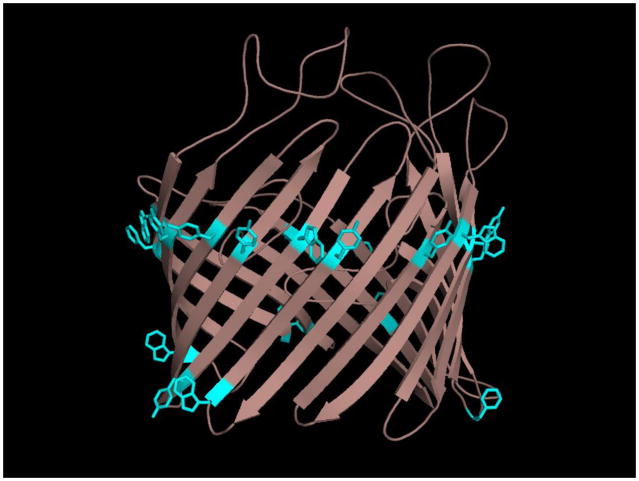
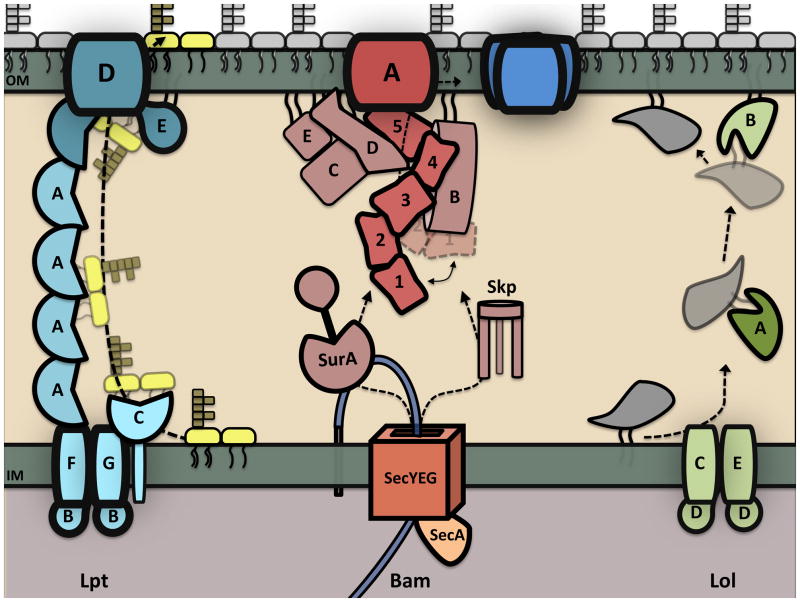
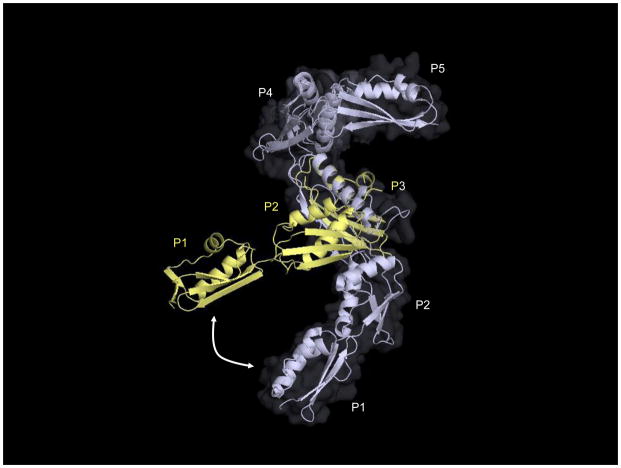
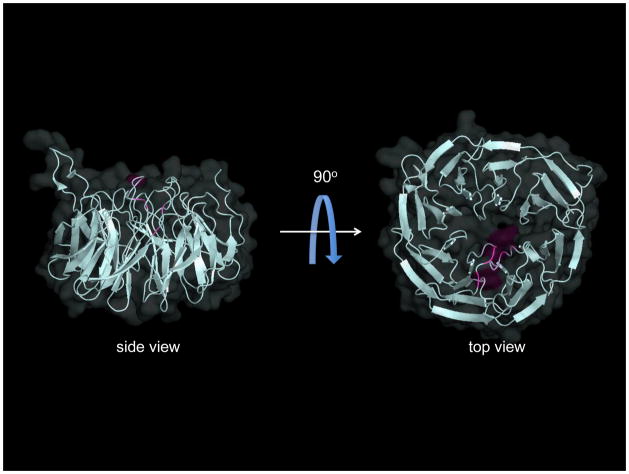
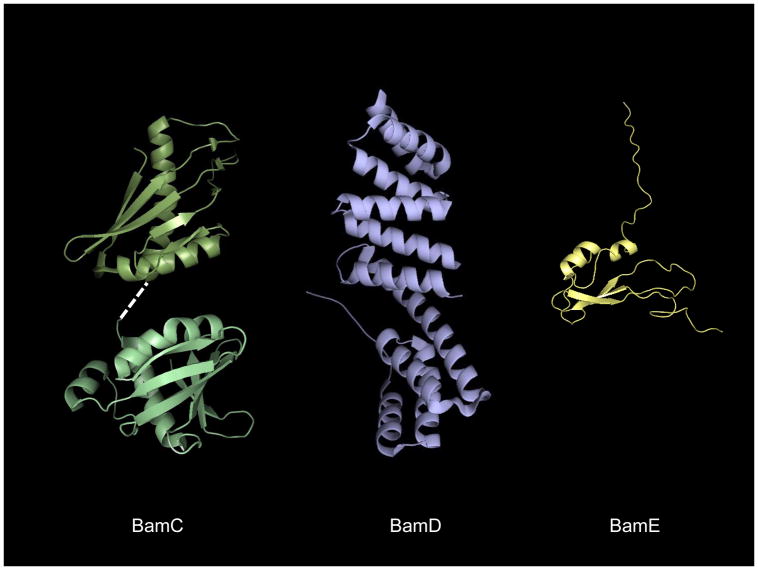
References
-
- Gray MW, Burger G, Lang BF. Mitochondrial evolution. Science. 1999;283:1476–1481. - PubMed
-
- Lugtenberg EJ, Peters R. Distribution of lipids in cytoplasmic and outer membranes of Escherichia coli K12. Biochim Biophys Acta. 1976;441:38–47. - PubMed
-
- Huijbregts RP, de Kroon A, de Kruijff B. Topology and transport of membrane lipids in bacteria. Biochim Biophys Acta. 2000;1469:43–61. - PubMed
-
- Kamio Y, Nikaido H. Outer membrane of Salmonella typhimurium: accessibility of phospholipid head groups to phospholipase c and cyanogen bromide activated dextran in the external medium. Biochemistry. 1976;15:2561–2570. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
