Multiple arrhythmic syndromes in a newborn, owing to a novel mutation in SCN5A
- PMID: 21895525
- PMCID: PMC3208331
- DOI: 10.1139/y11-070
Multiple arrhythmic syndromes in a newborn, owing to a novel mutation in SCN5A
Abstract
Background: Mutations in the SCN5A gene have been linked to Brugada syndrome (BrS), conduction disease, Long QT syndrome (LQT3), atrial fibrillation (AF), and to pre- and neonatal ventricular arrhythmias.
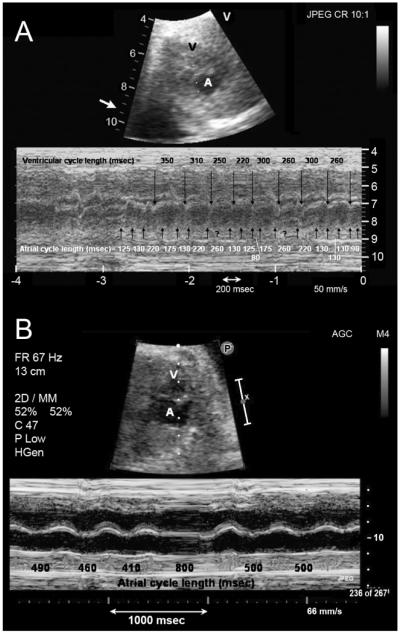
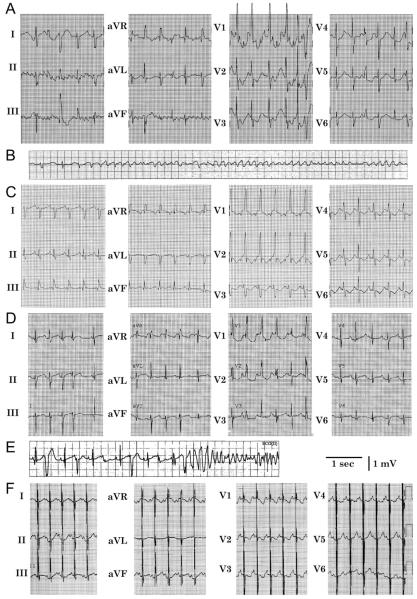
Objective: The objective of this study is to characterize a novel mutation in Na(v)1.5 found in a newborn with fetal chaotic atrial tachycardia, post-partum intraventricular conduction delay, and QT interval prolongation.
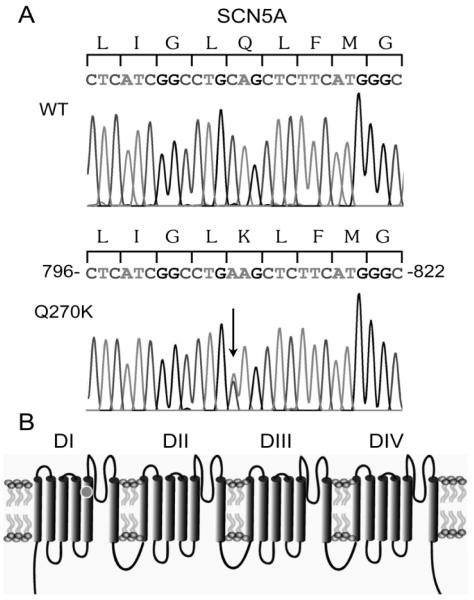
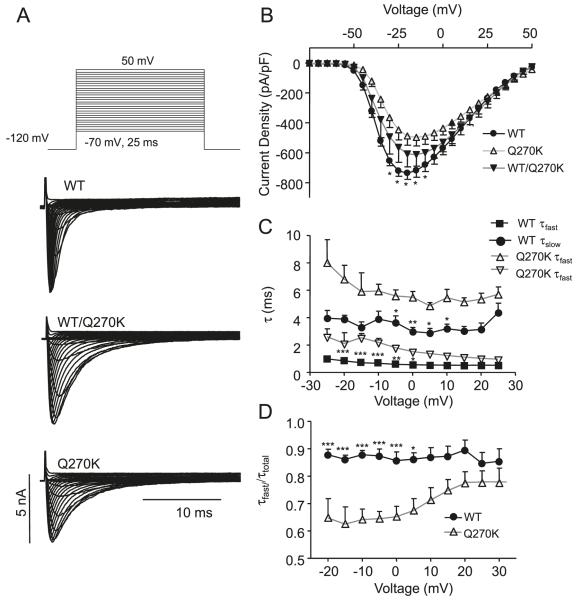
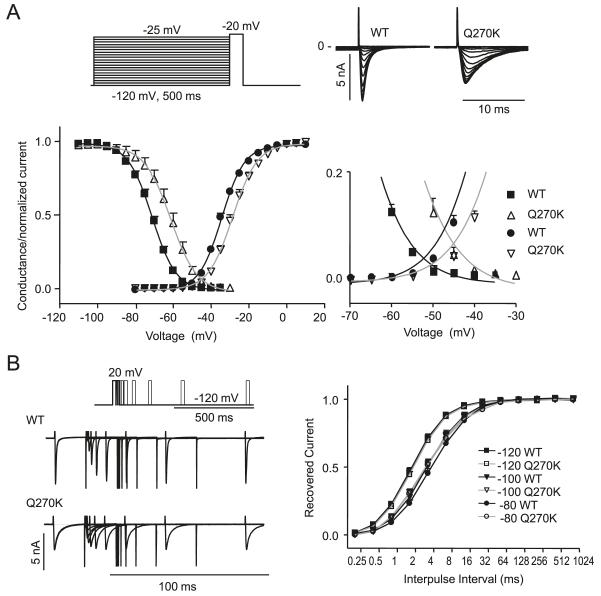
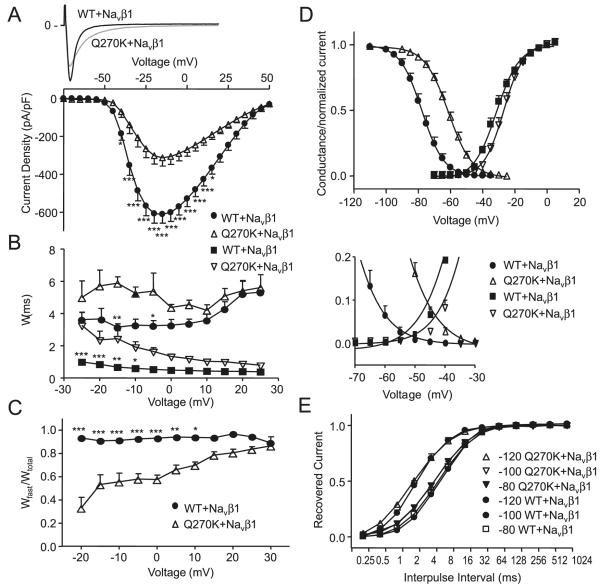
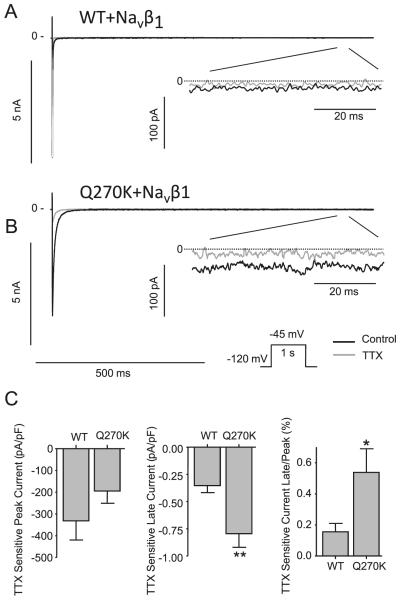
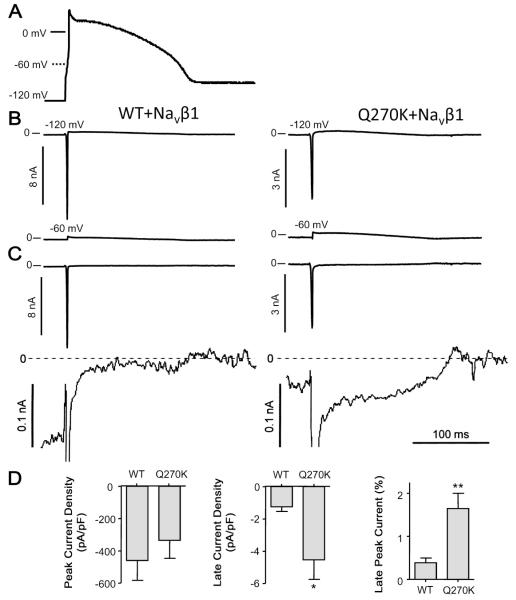
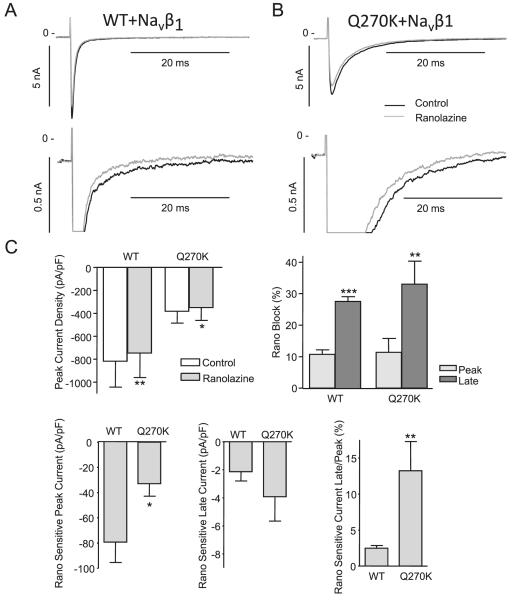
Methods: Genomic DNA was isolated and all exons and intron borders of 15 ion-channel genes were sequenced, revealing a novel missense mutation (Q270K) in SCN5A. Na(v)1.5 wild type (WT) and Q270K were expressed in CHO-K1 with and without the Na(v)β1 subunit. Results. Patch-clamp analysis showed ∼40% reduction in peak sodium channel current (I(Na)) density for Q270K compared with WT. Fast and slow decay of I(Na) were significantly slower in Q270K. Steady-state activation and inactivation of Q270K channels were shifted to positive potentials, and window current was increased. The tetrodotoxin-sensitive late I(Na) was increased almost 3-fold compared with WT channels. Ranolazine reduced late I(Na) in WT and Q270K channels, while exerting minimal effects on peak I(Na).
Conclusion: The Q270K mutation in SCN5A reduces peak I(Na) while augmenting late I(Na), and may thus underlie the development of atrial tachycardia, intraventricular conduction delay, and QT interval prolongation in an infant.
Figures
References
-
- Abriel H. Cardiac sodium channel Na(v)1.5 and interacting proteins: physiology and pathophysiology. J. Mol. Cell. Cardiol. 2010;48(1):2–11. doi:10.1016/j.yjmcc.2009.08.025. PMID:19744495. - PubMed
-
- Ackerman MJ, Siu BL, Sturner WQ, Tester DJ, Valdivia CR, Makielski JC, Towbin JA. Postmortem molecular analysis of SCN5A defects in sudden infant death syndrome. JAMA. 2001;286(18):2264–2269. doi:10.1001/jama.286.18.2264. PMID:11710892. - PubMed
-
- Antzelevitch C, Brugada P, Borggrefe M, Brugada J, Brugada R, Corrado D, et al. Brugada syndrome: report of the second consensus conference. Endorsed by the Heart Rhythm Society and the European Heart Rhythm Association. Circulation. 2005;111(5):659–670. doi:10.1161/01.CIR.0000152479.54298.51. PMID:15655131. - PubMed
-
- Arnestad M, Crotti L, Rognum TO, Insolia R, Pedrazzini M, Ferrandi C, et al. Prevalence of long-QT syndrome gene variants in sudden infant death syndrome. Circulation. 2007;115(3):361–367. doi:10.1161/CIRCULATIONAHA.106.658021. PMID: 17210839. - PubMed
-
- Attwell D, Cohen I, Eisner D, Ohba M, Ojeda C. The steady state TTX-sensitive ("window") sodium current in cardiac Purkinje fibres. Pflugers Arch. 1979;379(2):137–142. doi:10.1007/ BF00586939. PMID:571107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous