

The endosomal Na(+)/H(+) exchanger contributes to multivesicular body formation by regulating the recruitment of ESCRT-0 Vps27p to the endosomal membrane
- PMID: 21896492
- PMCID: PMC3199507
- DOI: 10.1074/jbc.M111.260612
The endosomal Na(+)/H(+) exchanger contributes to multivesicular body formation by regulating the recruitment of ESCRT-0 Vps27p to the endosomal membrane
Abstract
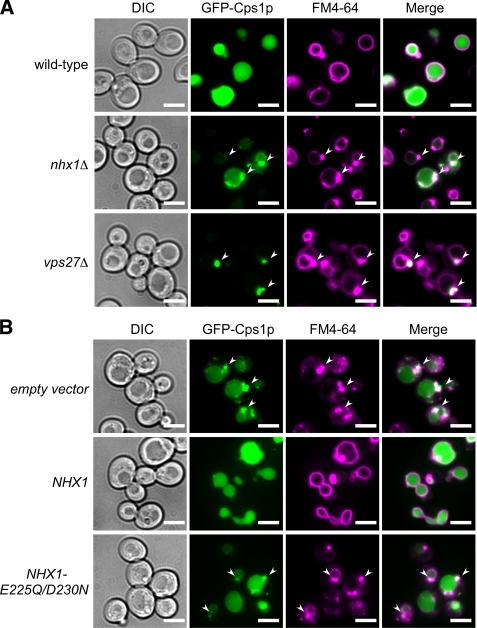
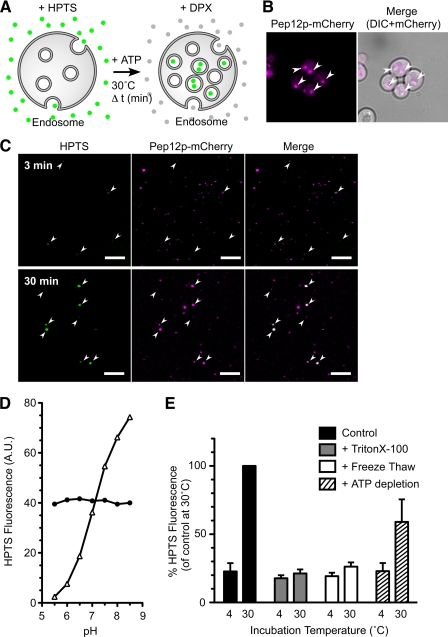
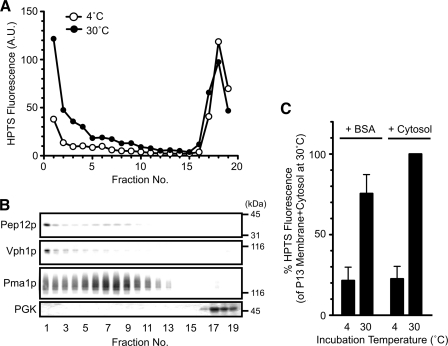
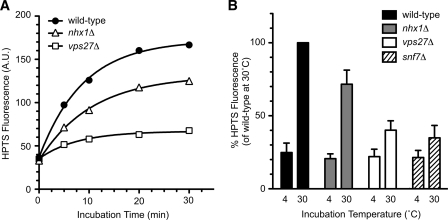
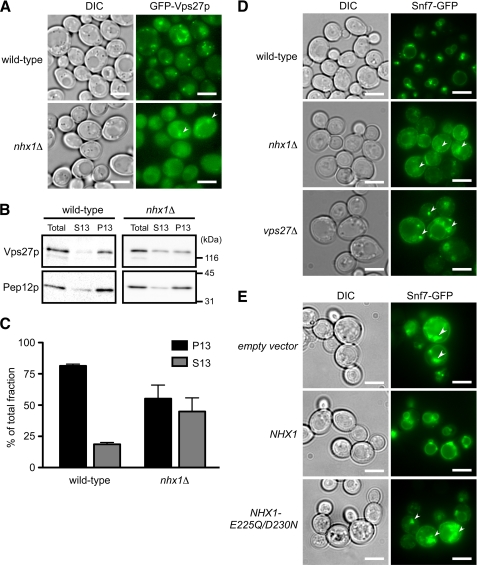
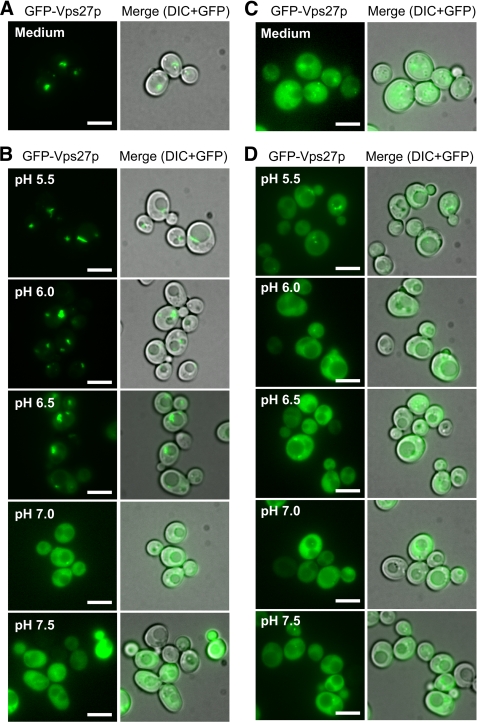
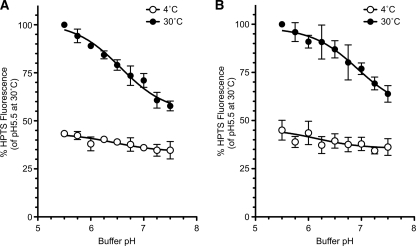
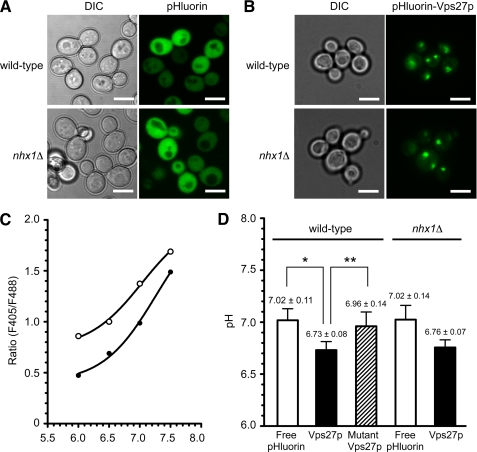
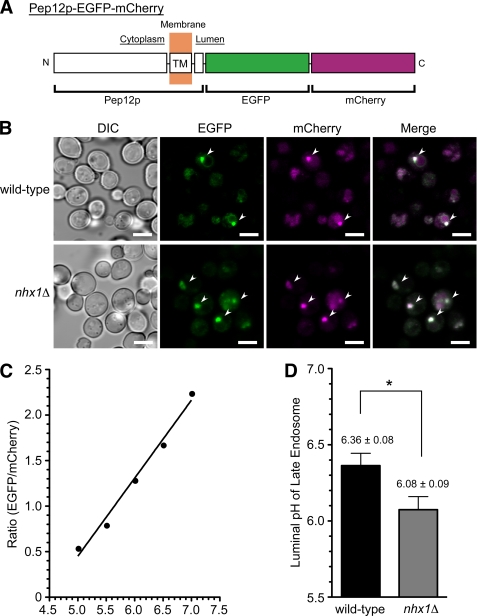
Multivesicular bodies (MVBs) are late endosomal compartments containing luminal vesicles (MVB vesicles) that are formed by inward budding of the endosomal membrane. In budding yeast, MVBs are an important cellular mechanism for the transport of membrane proteins to the vacuolar lumen. This process requires a class E subset of vacuolar protein sorting (VPS) genes. VPS44 (allelic to NHX1) encodes an endosome-localized Na(+)/H(+) exchanger. The function of the VPS44 exchanger in the context of vacuolar protein transport is largely unknown. Using a cell-free MVB formation assay system, we demonstrated that Nhx1p is required for the efficient formation of MVB vesicles in the late endosome. The recruitment of Vps27p, a class E Vps protein, to the endosomal membrane was dependent on Nhx1p activity and was enhanced by an acidic pH at the endosomal surface. Taken together, we propose that Nhx1p contributes to MVB formation by the recruitment of Vps27p to the endosomal membrane, possibly through Nhx1p antiporter activity.
Figures
References
-
- Lee J. A., Beigneux A., Ahmad S. T., Young S. G., Gao F. B. (2007) Curr. Biol. 17, 1561–1567 - PubMed
-
- Katzmann D. J., Odorizzi G., Emr S. D. (2002) Nat. Rev. Mol. Cell Biol. 3, 893–905 - PubMed
-
- Raiborg C., Stenmark H. (2009) Nature 458, 445–452 - PubMed
-
- Carlton J. G., Martin-Serrano J. (2007) Science 316, 1908–1912 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
