

Ending the message: poly(A) signals then and now
- PMID: 21896654
- PMCID: PMC3175714
- DOI: 10.1101/gad.17268411
Ending the message: poly(A) signals then and now
Abstract
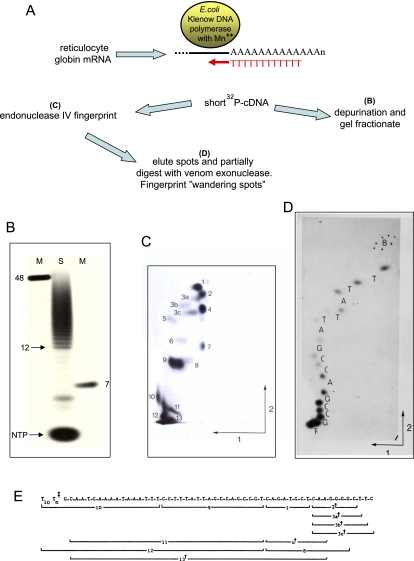
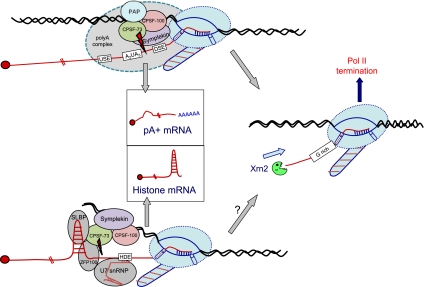
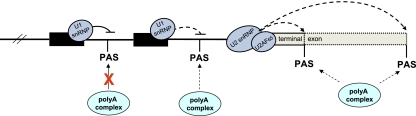
Polyadenylation [poly(A)] signals (PAS) are a defining feature of eukaryotic protein-coding genes. The central sequence motif AAUAAA was identified in the mid-1970s and subsequently shown to require flanking, auxiliary elements for both 3'-end cleavage and polyadenylation of premessenger RNA (pre-mRNA) as well as to promote downstream transcriptional termination. More recent genomic analysis has established the generality of the PAS for eukaryotic mRNA. Evidence for the mechanism of mRNA 3'-end formation is outlined, as is the way this RNA processing reaction communicates with RNA polymerase II to terminate transcription. The widespread phenomenon of alternative poly(A) site usage and how this interrelates with pre-mRNA splicing is then reviewed. This shows that gene expression can be drastically affected by how the message is ended. A central theme of this review is that while genomic analysis provides generality for the importance of PAS selection, detailed mechanistic understanding still requires the direct analysis of specific genes by genetic and biochemical approaches.
Figures

References
-
- Adesnik M, Salditt M, Thomas W, Darnell JE 1972. Evidence that all messenger RNA molecules (except histone messenger RNA) contain poly (A) sequences and that the poly(A) has a nuclear function. J Mol Biol 71: 21–30 - PubMed
-
- Amara SG, Jonas V, Rosenfeld MG, Ong ES, Evans RM 1982. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature 298: 240–244 - PubMed
-
- Ashe MP, Griffin P, James W, Proudfoot NJ 1995. Poly(A) site selection in the HIV-1 provirus: inhibition of promoter-proximal polyadenylation by the downstream major splice donor site. Genes Dev 9: 3008–3025 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources