

Structural basis for the role of the Sir3 AAA+ domain in silencing: interaction with Sir4 and unmethylated histone H3K79
- PMID: 21896656
- PMCID: PMC3175719
- DOI: 10.1101/gad.17175111
Structural basis for the role of the Sir3 AAA+ domain in silencing: interaction with Sir4 and unmethylated histone H3K79
Abstract
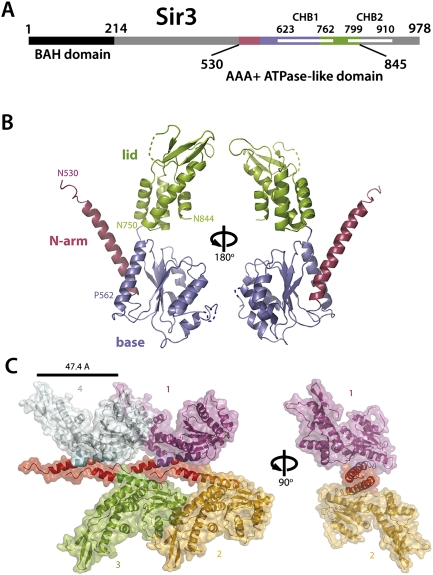
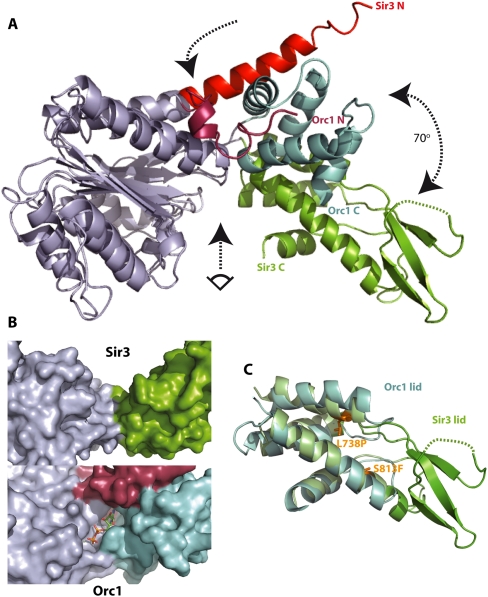
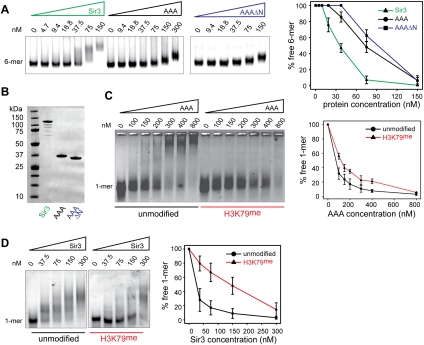
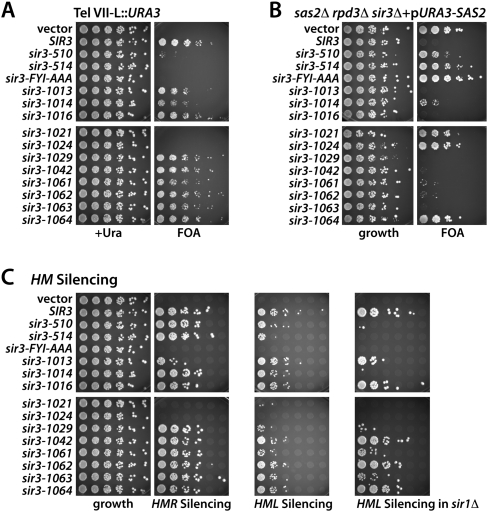
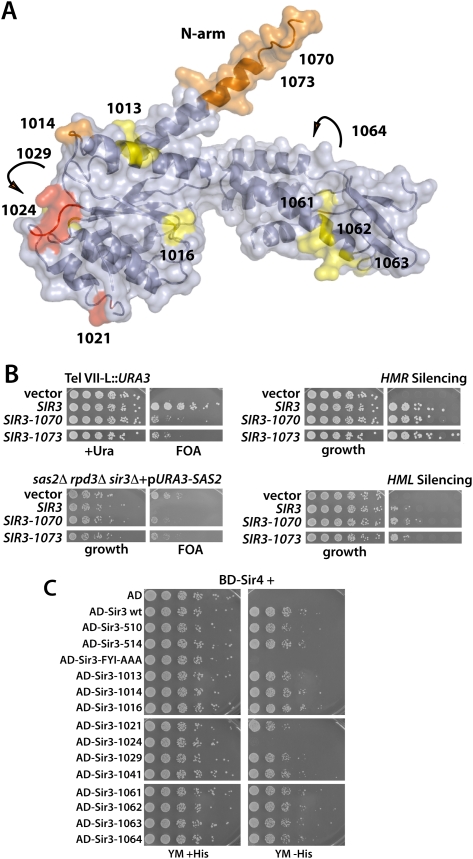
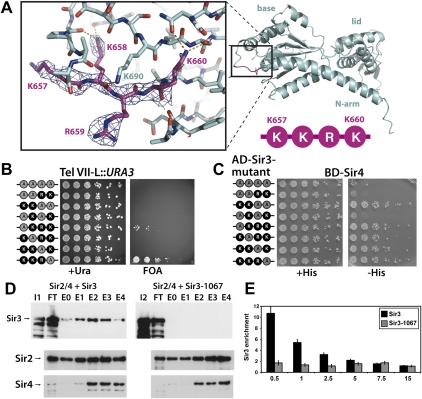
The silent information regulator 2/3/4 (Sir2/3/4) complex is required for gene silencing at the silent mating-type loci and at telomeres in Saccharomyces cerevisiae. Sir3 is closely related to the origin recognition complex 1 subunit and consists of an N-terminal bromo-adjacent homology (BAH) domain and a C-terminal AAA(+) ATPase-like domain. Here, through a combination of structure biology and exhaustive mutagenesis, we identified unusual, silencing-specific features of the AAA(+) domain of Sir3. Structural analysis of the putative nucleotide-binding pocket in this domain reveals a shallow groove that would preclude nucleotide binding. Mutation of this site has little effect on Sir3 function in vivo. In contrast, several surface regions are shown to be necessary for the Sir3 silencing function. Interestingly, the Sir3 AAA(+) domain is shown here to bind chromatin in vitro in a manner sensitive to histone H3K79 methylation. Moreover, an exposed loop on the surface of this Sir3 domain is found to interact with Sir4. In summary, the unique folding of this conserved Sir3 AAA(+) domain generates novel surface regions that mediate Sir3-Sir4 and Sir3-nucleosome interactions, both being required for the proper assembly of heterochromatin in living cells.
Figures
References
-
- Bell SP, Mitchell J, Leber J, Kobayashi R, Stillman B 1995. The multidomain structure of Orc1p reveals similarity to regulators of DNA replication and transcriptional silencing. Cell 83: 563–568 - PubMed
-
- Braunstein M, Rose AB, Holmes SG, Allis CD, Broach JR 1993. Transcriptional silencing in yeast is associated with reduced nucleosome acetylation. Genes Dev 7: 592–604 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases