

Human SNM1A and XPF-ERCC1 collaborate to initiate DNA interstrand cross-link repair
- PMID: 21896658
- PMCID: PMC3175721
- DOI: 10.1101/gad.15699211
Human SNM1A and XPF-ERCC1 collaborate to initiate DNA interstrand cross-link repair
Abstract
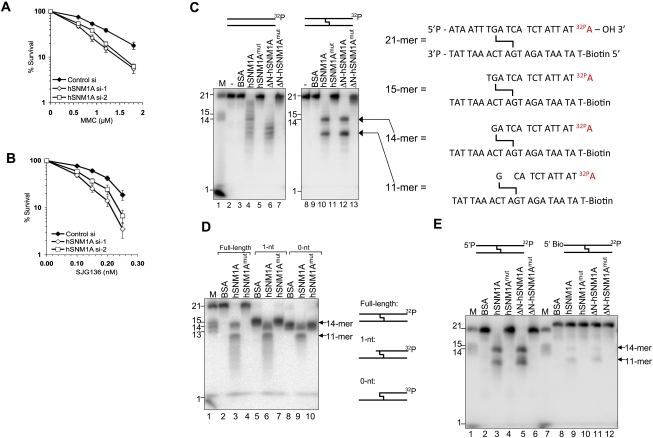
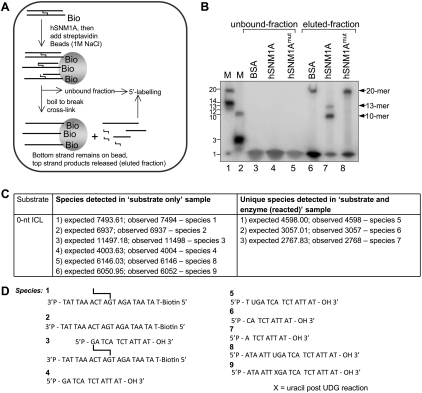
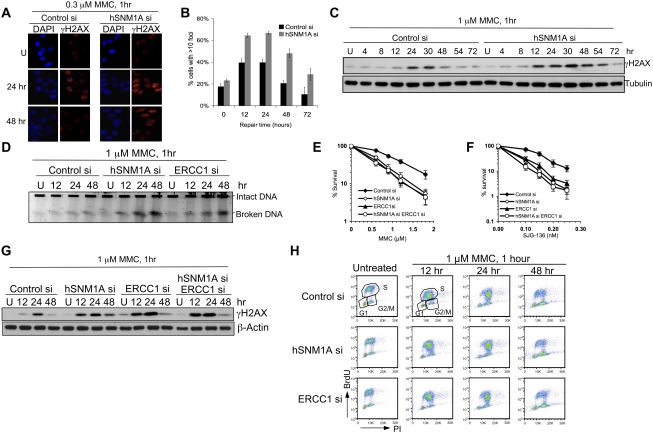
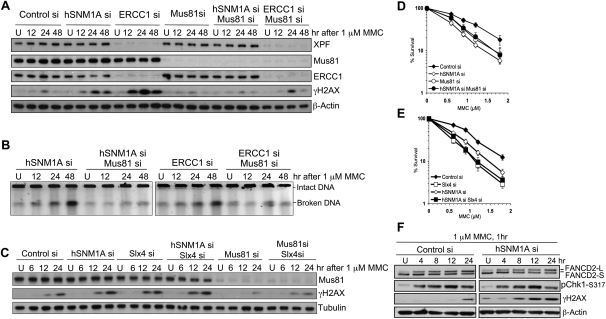
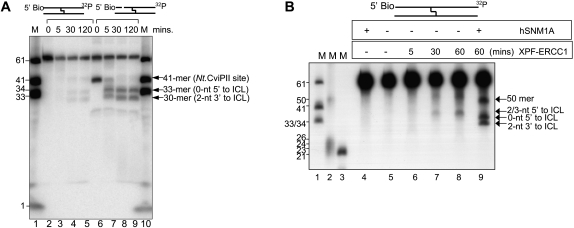
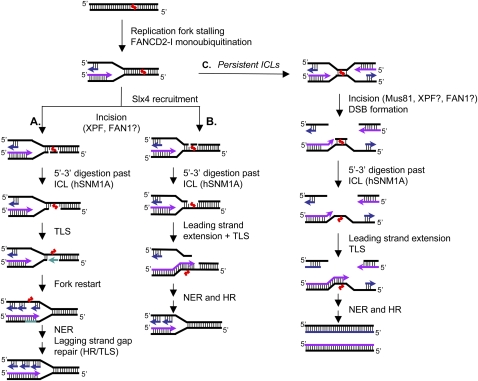
One of the major DNA interstrand cross-link (ICL) repair pathways in mammalian cells is coupled to replication, but the mechanistic roles of the critical factors involved remain largely elusive. Here, we show that purified human SNM1A (hSNM1A), which exhibits a 5'-3' exonuclease activity, can load from a single DNA nick and digest past an ICL on its substrate strand. hSNM1A-depleted cells are ICL-sensitive and accumulate replication-associated DNA double-strand breaks (DSBs), akin to ERCC1-depleted cells. These DSBs are Mus81-induced, indicating that replication fork cleavage by Mus81 results from the failure of the hSNM1A- and XPF-ERCC1-dependent ICL repair pathway. Our results reveal how collaboration between hSNM1A and XPF-ERCC1 is necessary to initiate ICL repair in replicating human cells.
Figures
Similar articles
-
RPA activates the XPF-ERCC1 endonuclease to initiate processing of DNA interstrand crosslinks.EMBO J. 2017 Jul 14;36(14):2047-2060. doi: 10.15252/embj.201796664. Epub 2017 Jun 12. EMBO J. 2017. PMID: 28607004 Free PMC article.
-
Orchestrating the nucleases involved in DNA interstrand cross-link (ICL) repair.Cell Cycle. 2011 Dec 1;10(23):3999-4008. doi: 10.4161/cc.10.23.18385. Epub 2011 Dec 1. Cell Cycle. 2011. PMID: 22101340 Free PMC article. Review.
-
XPF-ERCC1 participates in the Fanconi anemia pathway of cross-link repair.Mol Cell Biol. 2009 Dec;29(24):6427-37. doi: 10.1128/MCB.00086-09. Epub 2009 Oct 5. Mol Cell Biol. 2009. PMID: 19805513 Free PMC article.
-
The structure-specific endonuclease Ercc1-Xpf is required to resolve DNA interstrand cross-link-induced double-strand breaks.Mol Cell Biol. 2004 Jul;24(13):5776-87. doi: 10.1128/MCB.24.13.5776-5787.2004. Mol Cell Biol. 2004. PMID: 15199134 Free PMC article.
-
Interstrand crosslink repair: can XPF-ERCC1 be let off the hook?Trends Genet. 2008 Feb;24(2):70-6. doi: 10.1016/j.tig.2007.11.003. Epub 2008 Jan 14. Trends Genet. 2008. PMID: 18192062 Review.
Cited by
-
FANCD2-associated nuclease 1, but not exonuclease 1 or flap endonuclease 1, is able to unhook DNA interstrand cross-links in vitro.J Biol Chem. 2015 Sep 11;290(37):22602-11. doi: 10.1074/jbc.M115.663666. Epub 2015 Jul 28. J Biol Chem. 2015. PMID: 26221031 Free PMC article.
-
Polymorphisms in ERCC1 and XPF genes and risk of gastric cancer in an eastern Chinese population.PLoS One. 2012;7(11):e49308. doi: 10.1371/journal.pone.0049308. Epub 2012 Nov 15. PLoS One. 2012. PMID: 23166636 Free PMC article.
-
Navigating oxygen deprivation: liver transcriptomic responses of the red eared slider turtle to environmental anoxia.PeerJ. 2019 Nov 26;7:e8144. doi: 10.7717/peerj.8144. eCollection 2019. PeerJ. 2019. PMID: 31788367 Free PMC article.
-
DNA repair, recombination, and damage signaling.Genetics. 2022 Feb 4;220(2):iyab178. doi: 10.1093/genetics/iyab178. Genetics. 2022. PMID: 35137093 Free PMC article. Review.
-
The yeast Hrq1 helicase stimulates Pso2 translesion nuclease activity and thereby promotes DNA interstrand crosslink repair.J Biol Chem. 2020 Jul 3;295(27):8945-8957. doi: 10.1074/jbc.RA120.013626. Epub 2020 May 5. J Biol Chem. 2020. PMID: 32371399 Free PMC article.
References
-
- Andersson BS, Sadeghi T, Siciliano MJ, Legerski R, Murray D 1996. Nucleotide excision repair genes as determinants of cellular sensitivity to cyclophosphamide analogs. Cancer Chemother Pharmacol 38: 406–416 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous