

Induction of the alternative NF-κB pathway by lymphotoxin αβ (LTαβ) relies on internalization of LTβ receptor
- PMID: 21896778
- PMCID: PMC3209329
- DOI: 10.1128/MCB.05033-11
Induction of the alternative NF-κB pathway by lymphotoxin αβ (LTαβ) relies on internalization of LTβ receptor
Abstract
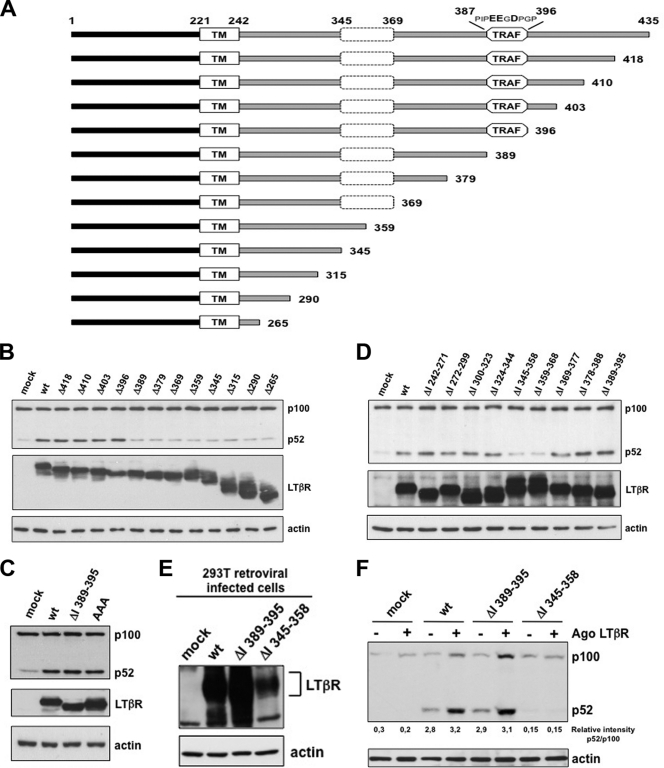
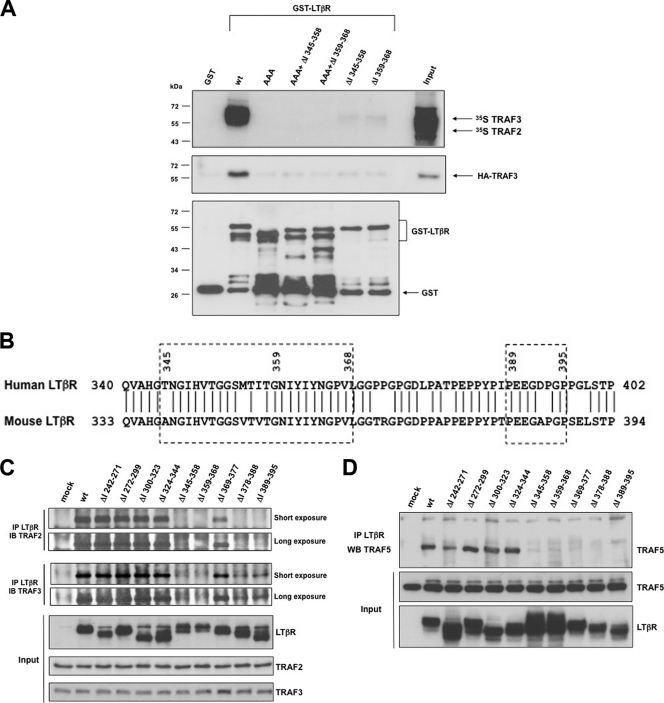
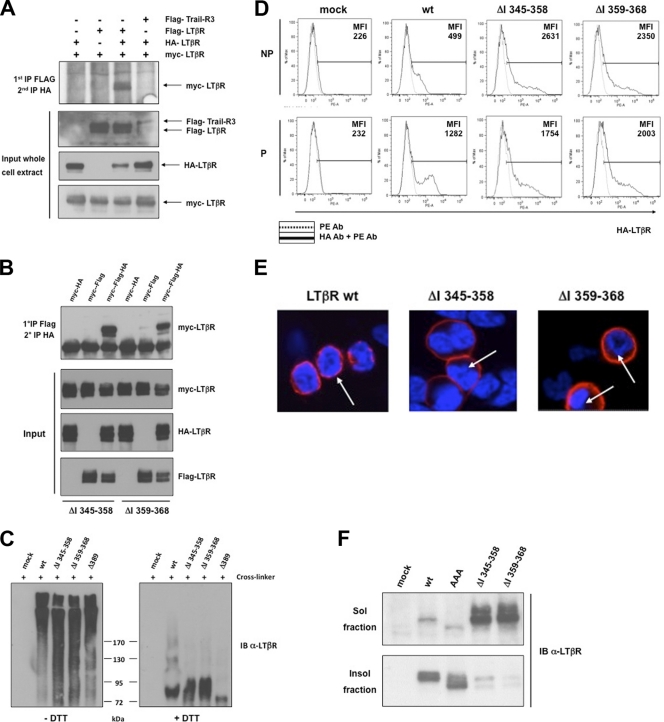
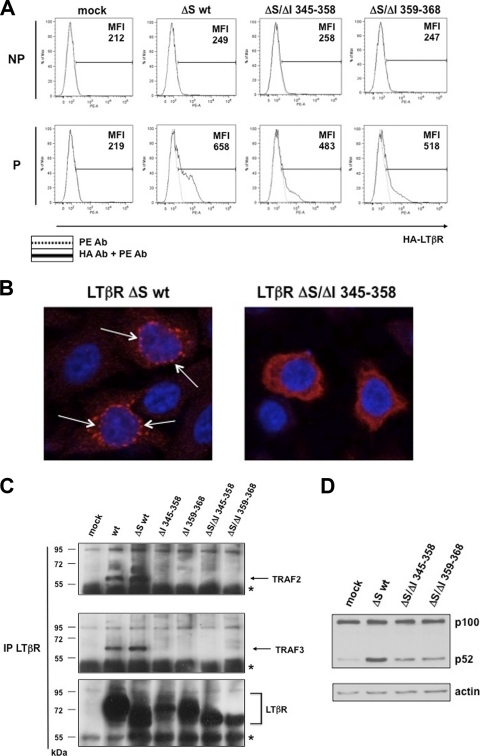
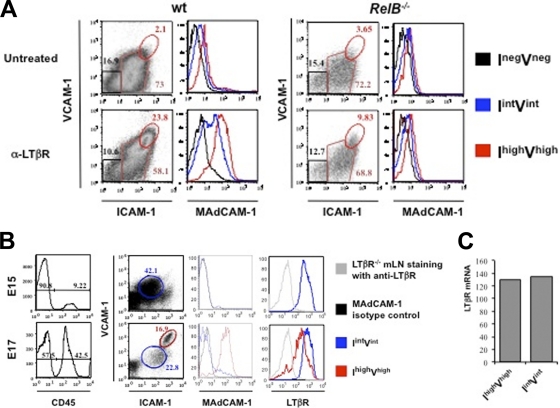
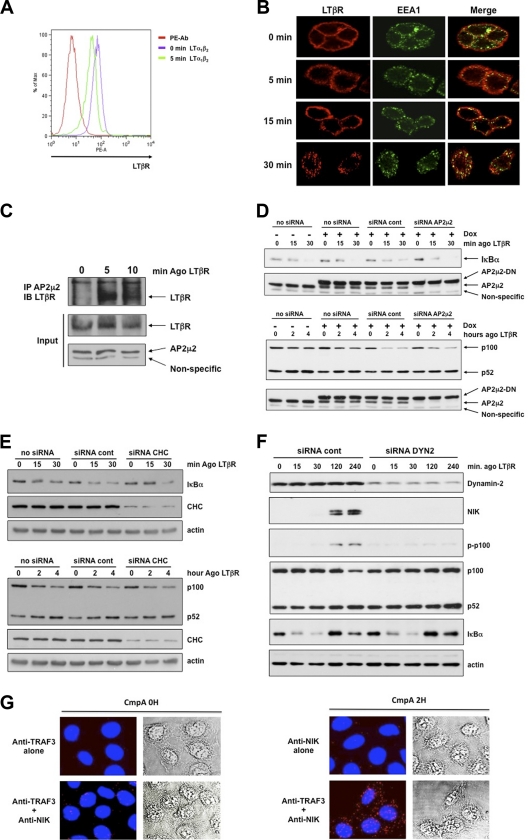
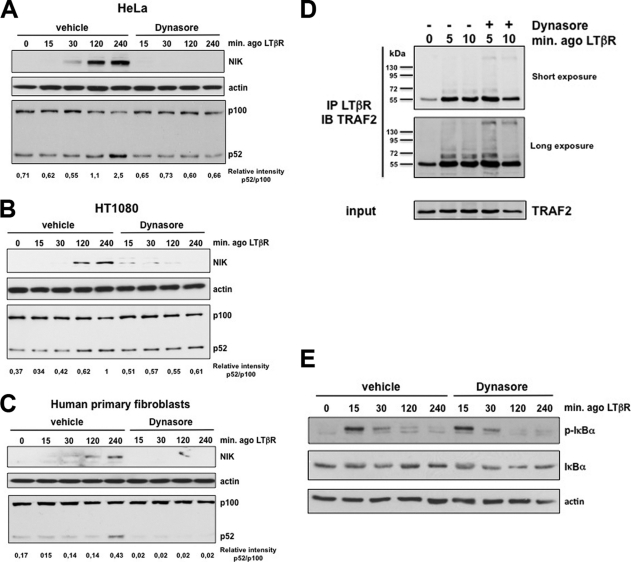
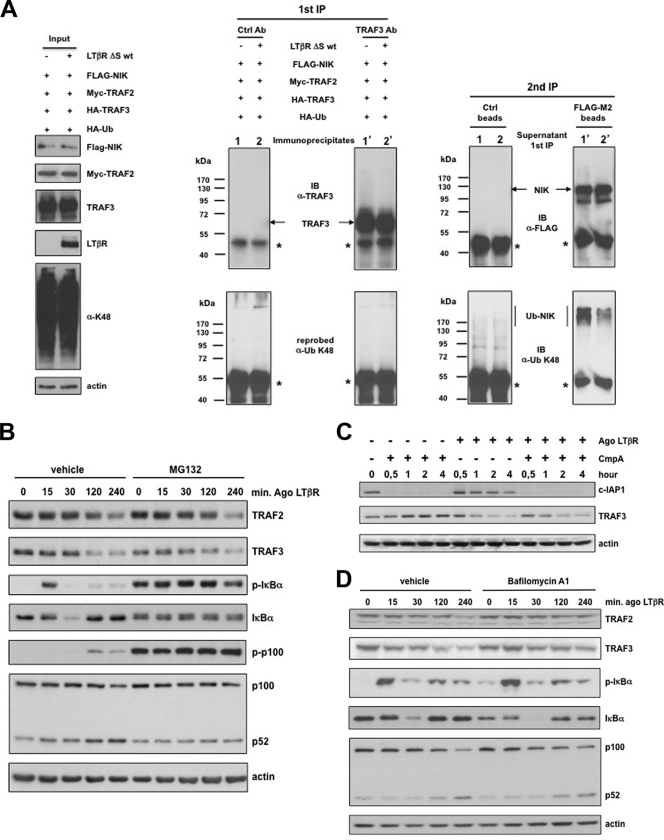
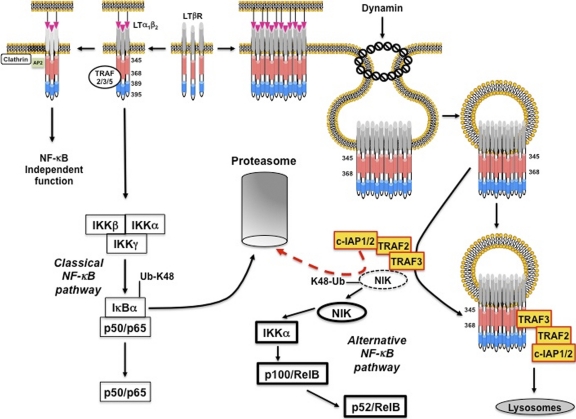
Several tumor necrosis factor receptor (TNFR) family members activate both the classical and the alternative NF-κB pathways. However, how a single receptor engages these two distinct pathways is still poorly understood. Using lymphotoxin β receptor (LTβR) as a prototype, we showed that activation of the alternative, but not the classical, NF-κB pathway relied on internalization of the receptor. Further molecular analyses revealed a specific cytosolic region of LTβR essential for its internalization, TRAF3 recruitment, and p100 processing. Interestingly, we found that dynamin-dependent, but clathrin-independent, internalization of LTβR appeared to be required for the activation of the alternative, but not the classical, NF-κB pathway. In vivo, ligand-induced internalization of LTβR in mesenteric lymph node stromal cells correlated with induction of alternative NF-κB target genes. Thus, our data shed light on LTβR cellular trafficking as a process required for specific biological functions of NF-κB.
Figures
References
-
- Batten M., et al. 2004. TNF deficiency fails to protect BAFF transgenic mice against autoimmunity and reveals a predisposition to B cell lymphoma. J. Immunol. 172:812–822 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials