

Chromosomes carrying meiotic avoidance loci in three apomictic eudicot Hieracium subgenus Pilosella species share structural features with two monocot apomicts
- PMID: 21896890
- PMCID: PMC3252177
- DOI: 10.1104/pp.111.181164
Chromosomes carrying meiotic avoidance loci in three apomictic eudicot Hieracium subgenus Pilosella species share structural features with two monocot apomicts
Abstract
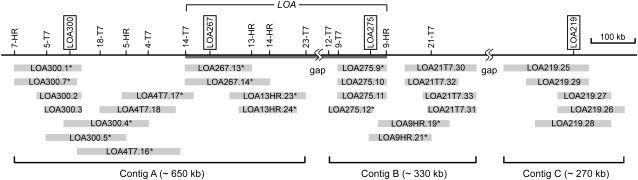
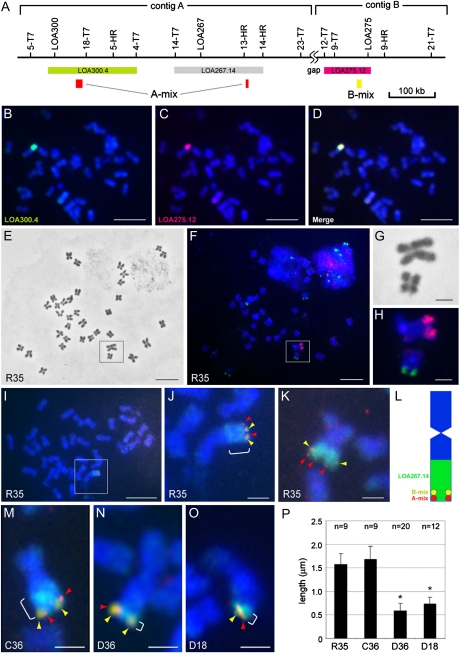
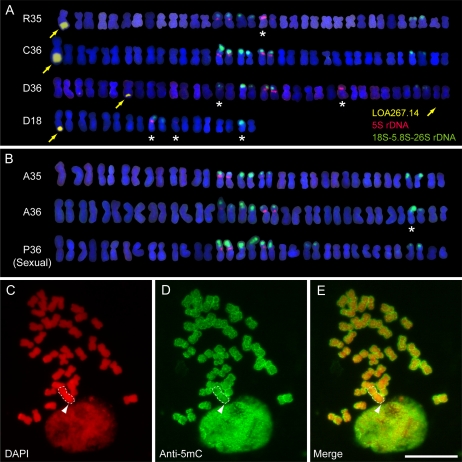
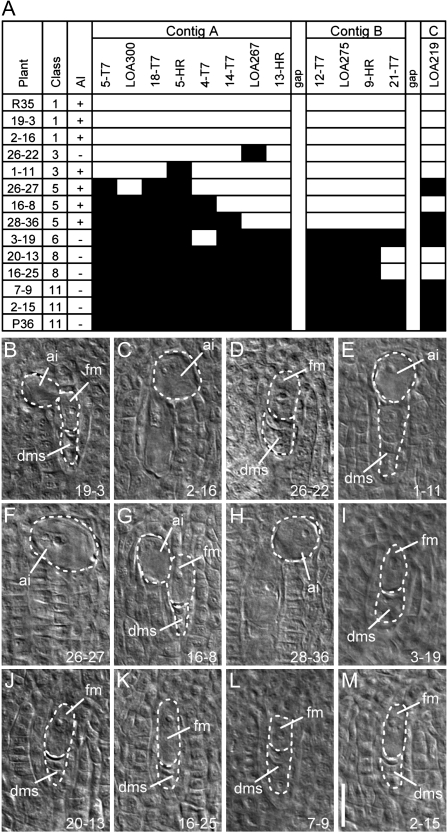
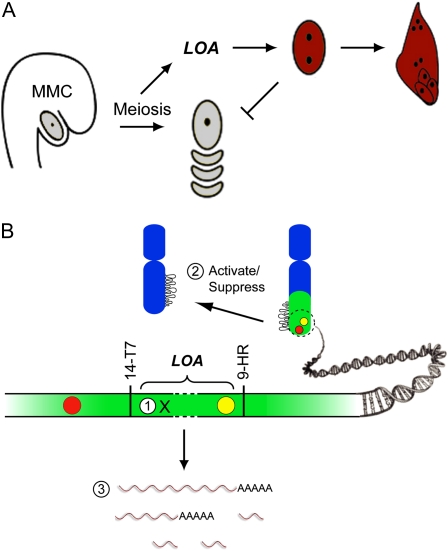
The LOSS OF APOMEIOSIS (LOA) locus is one of two dominant loci known to control apomixis in the eudicot Hieracium praealtum. LOA stimulates the differentiation of somatic aposporous initial cells after the initiation of meiosis in ovules. Aposporous initial cells undergo nuclear proliferation close to sexual megaspores, forming unreduced aposporous embryo sacs, and the sexual program ceases. LOA-linked genetic markers were used to isolate 1.2 Mb of LOA-associated DNAs from H. praealtum. Physical mapping defined the genomic region essential for LOA function between two markers, flanking 400 kb of identified sequence and central unknown sequences. Cytogenetic and sequence analyses revealed that the LOA locus is located on a single chromosome near the tip of the long arm and surrounded by extensive, abundant complex repeat and transposon sequences. Chromosomal features and LOA-linked markers are conserved in aposporous Hieracium caespitosum and Hieracium piloselloides but absent in sexual Hieracium pilosella. Their absence in apomictic Hieracium aurantiacum suggests that meiotic avoidance may have evolved independently in aposporous subgenus Pilosella species. The structure of the hemizygous chromosomal region containing the LOA locus in the three Hieracium subgenus Pilosella species resembles that of the hemizygous apospory-specific genomic regions in monocot Pennisetum squamulatum and Cenchrus ciliaris. Analyses of partial DNA sequences at these loci show no obvious conservation, indicating that they are unlikely to share a common ancestral origin. This suggests convergent evolution of repeat-rich hemizygous chromosomal regions containing apospory loci in these monocot and eudicot species, which may be required for the function and maintenance of the trait.
Figures
Similar articles
-
Evolution of apomixis loci in Pilosella and Hieracium (Asteraceae) inferred from the conservation of apomixis-linked markers in natural and experimental populations.Heredity (Edinb). 2015 Jan;114(1):17-26. doi: 10.1038/hdy.2014.61. Epub 2014 Jul 16. Heredity (Edinb). 2015. PMID: 25026970 Free PMC article.
-
A reference genetic linkage map of apomictic Hieracium species based on expressed markers derived from developing ovule transcripts.Ann Bot. 2015 Mar;115(4):567-80. doi: 10.1093/aob/mcu249. Epub 2014 Dec 22. Ann Bot. 2015. PMID: 25538115 Free PMC article.
-
The LOSS OF APOMEIOSIS (LOA) locus in Hieracium praealtum can function independently of the associated large-scale repetitive chromosomal structure.New Phytol. 2014 Feb;201(3):973-981. doi: 10.1111/nph.12574. Epub 2013 Nov 1. New Phytol. 2014. PMID: 24400904
-
Apomixis in hawkweed: Mendel's experimental nemesis.J Exp Bot. 2011 Mar;62(5):1699-707. doi: 10.1093/jxb/err011. Epub 2011 Feb 18. J Exp Bot. 2011. PMID: 21335438 Review.
-
Seeds of doubt: Mendel's choice of Hieracium to study inheritance, a case of right plant, wrong trait.Theor Appl Genet. 2016 Dec;129(12):2253-2266. doi: 10.1007/s00122-016-2788-x. Epub 2016 Oct 1. Theor Appl Genet. 2016. PMID: 27695890 Free PMC article. Review.
Cited by
-
Phenotypic plasticity of aposporous embryo sac development in Hieracium praealtum.Plant Signal Behav. 2019;14(8):1622981. doi: 10.1080/15592324.2019.1622981. Epub 2019 Jun 4. Plant Signal Behav. 2019. PMID: 31161899 Free PMC article.
-
Evolution of apomixis loci in Pilosella and Hieracium (Asteraceae) inferred from the conservation of apomixis-linked markers in natural and experimental populations.Heredity (Edinb). 2015 Jan;114(1):17-26. doi: 10.1038/hdy.2014.61. Epub 2014 Jul 16. Heredity (Edinb). 2015. PMID: 25026970 Free PMC article.
-
A reference genetic linkage map of apomictic Hieracium species based on expressed markers derived from developing ovule transcripts.Ann Bot. 2015 Mar;115(4):567-80. doi: 10.1093/aob/mcu249. Epub 2014 Dec 22. Ann Bot. 2015. PMID: 25538115 Free PMC article.
-
Generation of an integrated Hieracium genomic and transcriptomic resource enables exploration of small RNA pathways during apomixis initiation.BMC Biol. 2016 Oct 6;14(1):86. doi: 10.1186/s12915-016-0311-0. BMC Biol. 2016. PMID: 27716180 Free PMC article.
-
Genetic analyses of the inheritance and expressivity of autonomous endosperm formation in Hieracium with different modes of embryo sac and seed formation.Ann Bot. 2017 Apr 1;119(6):1001-1010. doi: 10.1093/aob/mcw262. Ann Bot. 2017. PMID: 28130222 Free PMC article.
References
-
- Akiyama Y, Hanna WW, Ozias-Akins P. (2005) High-resolution physical mapping reveals that the apospory-specific genomic region (ASGR) in Cenchrus ciliaris is located on a heterochromatic and hemizygous region of a single chromosome. Theor Appl Genet 111: 1042–1051 - PubMed
-
- Barreda VD, Palazzesi L, Tellería MC, Katinas L, Crisci JV, Bremer K, Passalia MG, Corsolini R, Rodríguez Brizuela R, Bechis F. (2010) Eocene Patagonia fossils of the daisy family. Science 329: 1621. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
