

Bivariate and Multivariate NeuroXidence: A Robust and Reliable Method to Detect Modulations of Spike-Spike Synchronization Across Experimental Conditions
- PMID: 21897816
- PMCID: PMC3158367
- DOI: 10.3389/fninf.2011.00014
Bivariate and Multivariate NeuroXidence: A Robust and Reliable Method to Detect Modulations of Spike-Spike Synchronization Across Experimental Conditions
Abstract
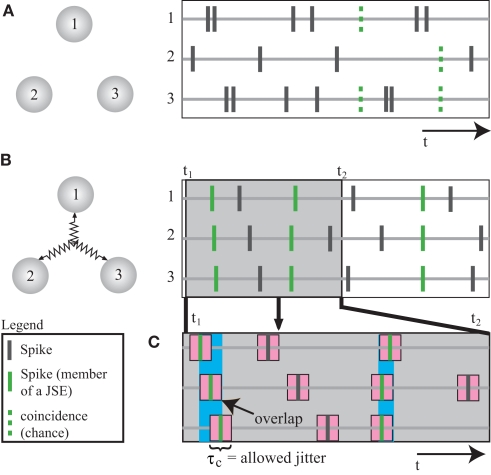
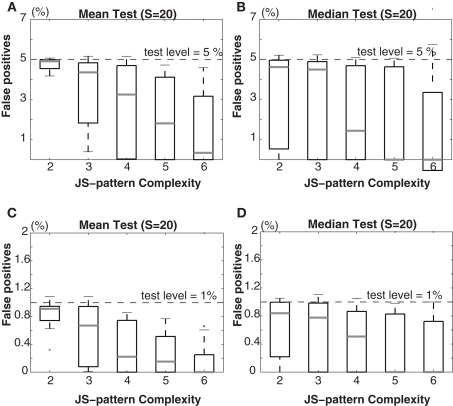
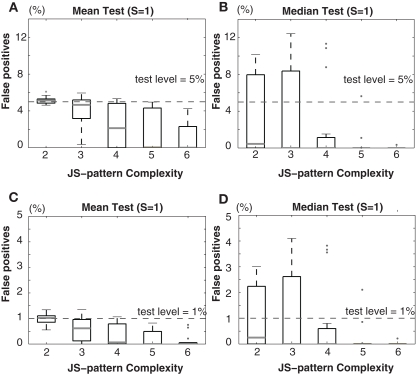
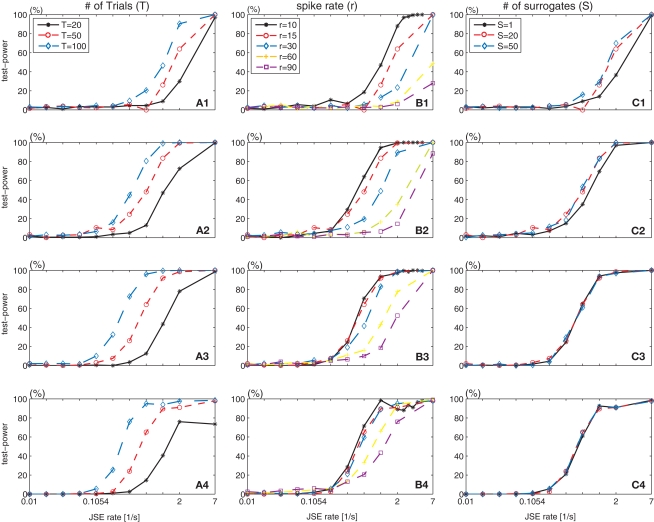
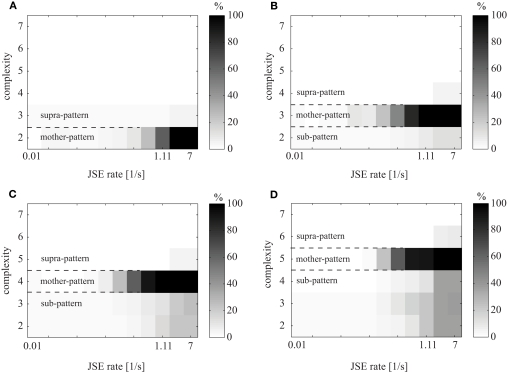
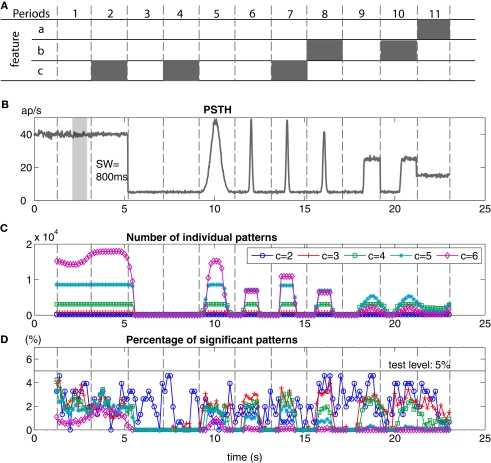
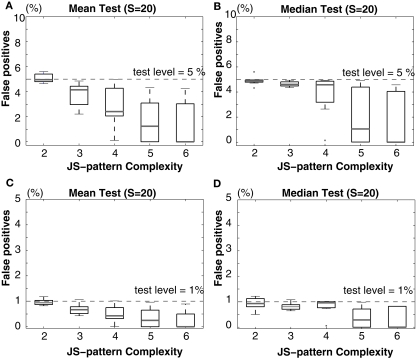
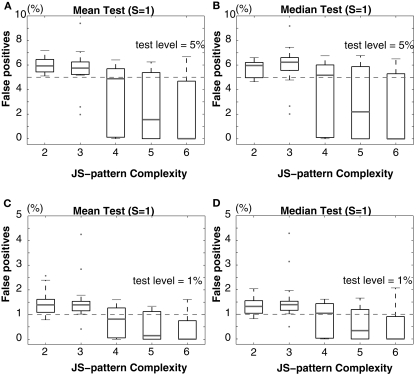
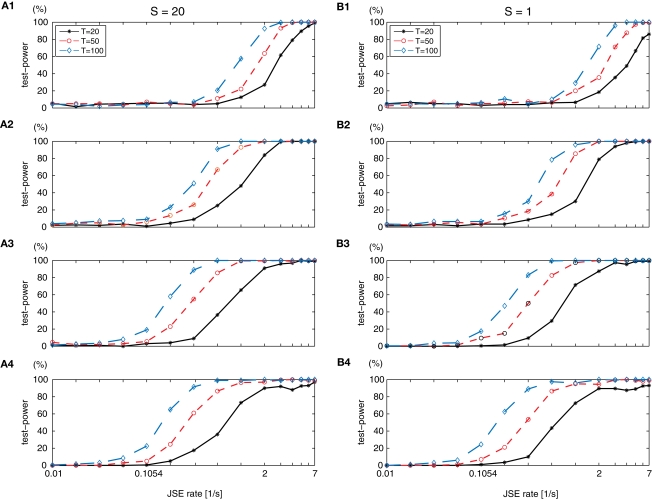
Synchronous neuronal firing has been proposed as a potential neuronal code. To determine whether synchronous firing is really involved in different forms of information processing, one needs to directly compare the amount of synchronous firing due to various factors, such as different experimental or behavioral conditions. In order to address this issue, we present an extended version of the previously published method, NeuroXidence. The improved method incorporates bi- and multivariate testing to determine whether different factors result in synchronous firing occurring above the chance level. We demonstrate through the use of simulated data sets that bi- and multivariate NeuroXidence reliably and robustly detects joint-spike-events across different factors.
Keywords: NeuroXidence; bivariate; factor; joint-spike-event; modulation of synchrony; multivariate; synchronous firing.
Figures
Similar articles
-
NeuroXidence: reliable and efficient analysis of an excess or deficiency of joint-spike events.J Comput Neurosci. 2008 Aug;25(1):64-88. doi: 10.1007/s10827-007-0065-3. Epub 2008 Jan 26. J Comput Neurosci. 2008. PMID: 18219568 Free PMC article.
-
Higher Order Spike Synchrony in Prefrontal Cortex during Visual Memory.Front Comput Neurosci. 2011 Jun 8;5:23. doi: 10.3389/fncom.2011.00023. eCollection 2011. Front Comput Neurosci. 2011. PMID: 21713065 Free PMC article.
-
Excess synchrony in motor cortical neurons provides redundant direction information with that from coarse temporal measures.J Neurophysiol. 2001 Oct;86(4):1700-16. doi: 10.1152/jn.2001.86.4.1700. J Neurophysiol. 2001. PMID: 11600633
-
A novel, jitter-based method for detecting and measuring spike synchrony and quantifying temporal firing precision.Neural Syst Circuits. 2012 May 2;2(1):5. doi: 10.1186/2042-1001-2-5. Neural Syst Circuits. 2012. PMID: 22551243 Free PMC article.
-
Diverse synchrony of firing reflects diverse cell-assembly coding in the prefrontal cortex.J Physiol Paris. 2013 Dec;107(6):459-70. doi: 10.1016/j.jphysparis.2013.05.004. Epub 2013 Jun 4. J Physiol Paris. 2013. PMID: 23747709 Review.
References
-
- Aertsen A., Diesmann M., Gewaltig M. O., Grün S., Rotter S. (2001). “Neural dynamics in cortical networks – precision of joint-spiking events,” in Comlexity in Biological Information Processing: Novartis Foundation Symposium, eds Bock G. R., Goode J. A. (Chichester: John Wiley & Sons, Ltd.), 239, 193–204 - PubMed
-
- Baker S. N., Lemon R. N. (2000). Precise spatiotemporal repeating patterns in monkey primary and supplementary motor areas occur at chance levels. J. Neurophysiol. 84, 1770–1780 - PubMed
LinkOut - more resources
Full Text Sources