

The ER in 3D: a multifunctional dynamic membrane network
- PMID: 21900009
- PMCID: PMC3221873
- DOI: 10.1016/j.tcb.2011.07.004
The ER in 3D: a multifunctional dynamic membrane network
Abstract
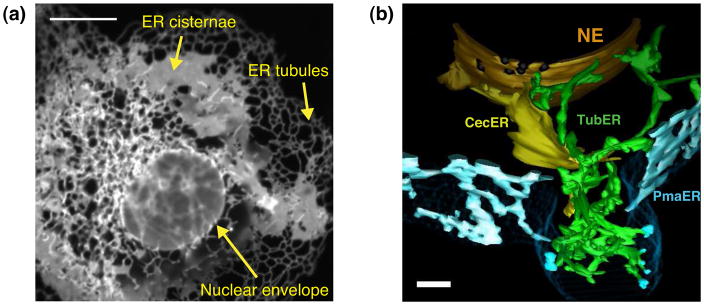
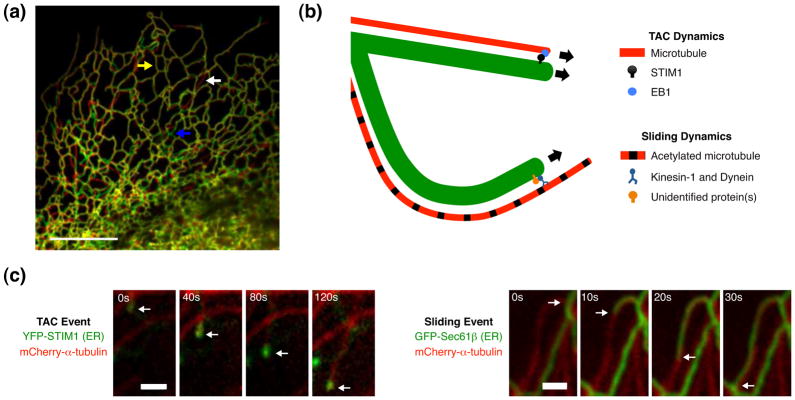
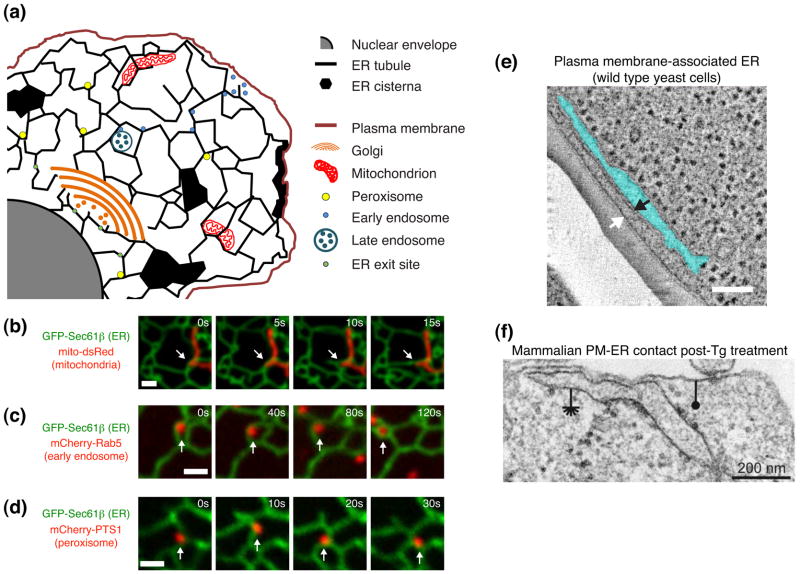
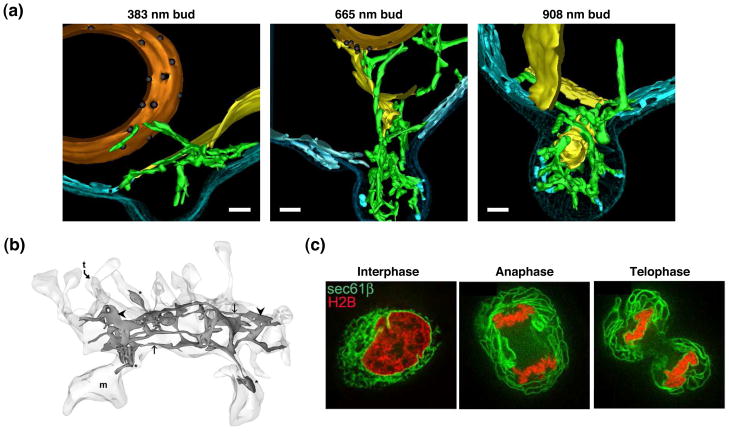
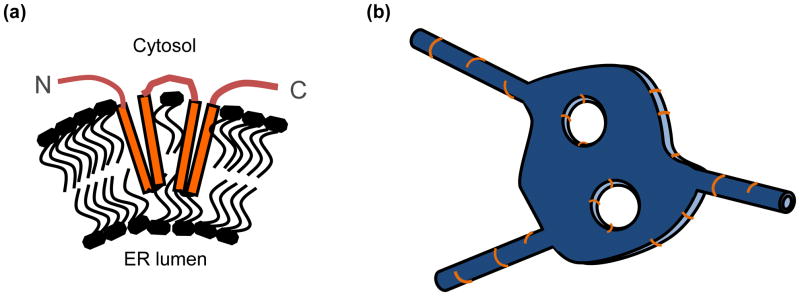
The endoplasmic reticulum (ER) is a large, singular, membrane-bound organelle that has an elaborate 3D structure with a diversity of structural domains. It contains regions that are flat and cisternal, ones that are highly curved and tubular, and others adapted to form contacts with nearly every other organelle and with the plasma membrane. The 3D structure of the ER is determined by both integral ER membrane proteins and by interactions with the cytoskeleton. In this review, we describe some of the factors that are known to regulate ER structure and discuss how this structural organization and the dynamic nature of the ER membrane network allow it to perform its many different functions.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Baumann O, Walz B. Endoplasmic reticulum of animal cells and its organization into structural and functional domains. Int Rev Cytol. 2001;205:149–214. - PubMed
-
- Hetzer MW, et al. Pushing the envelope: structure, function, and dynamics of the nuclear periphery. Annu Rev Cell Dev Biol. 2005;21:347–380. - PubMed
-
- Voeltz GK, et al. A class of membrane proteins shaping the tubular endoplasmic reticulum. Cell. 2006;124:573–586. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
