

Inhibition of IL-32 activation by α-1 antitrypsin suppresses alloreactivity and increases survival in an allogeneic murine marrow transplantation model
- PMID: 21900190
- PMCID: PMC3208308
- DOI: 10.1182/blood-2011-07-365247
Inhibition of IL-32 activation by α-1 antitrypsin suppresses alloreactivity and increases survival in an allogeneic murine marrow transplantation model
Abstract
Interleukin (IL)-32 was originally identified in natural killer cells and IL-2-activated human T lymphocytes. As T cells are activated in allogeneic transplantation, we determined the role of IL-32 in human mixed lymphocyte cultures (MLCs) and GVHD. In allogeneic MLCs, IL-32 increased two-fold in responding T cells, accompanied by five-fold increases of TNFα, IL-6, and IL-8. After allogeneic hematopoietic cell transplantation, IL-32 mRNA levels in blood leukocytes were statistically significantly higher in patients with acute GVHD (n = 10) than in serial samples from patients who did not develop acute GVHD (n = 5; P = .02). No significant changes in IL-32 levels were present in patients with treated (n = 14) or untreated (n = 8) chronic GVHD, compared with healthy controls (n = 8; P = .5, and P = .74, respectively). As IL-32 is activated by proteinase-3 (PR3), we determined the effect of the serine protease inhibitor α-1 antitrypsin (AAT) on IL-32 levels and showed suppression of IL-32 and T-lymphocyte proliferation in MLCs. In an MHC-minor antigen disparate murine transplant model, preconditioning and postconditioning treatment with AAT resulted in attenuation or prevention of GVHD and superior survival compared with albumin-treated controls (80% vs 44%; P = .04). These findings suggest that AAT modulates immune and inflammatory functions and may represent a novel approach to prevent or treat GVHD.
Figures


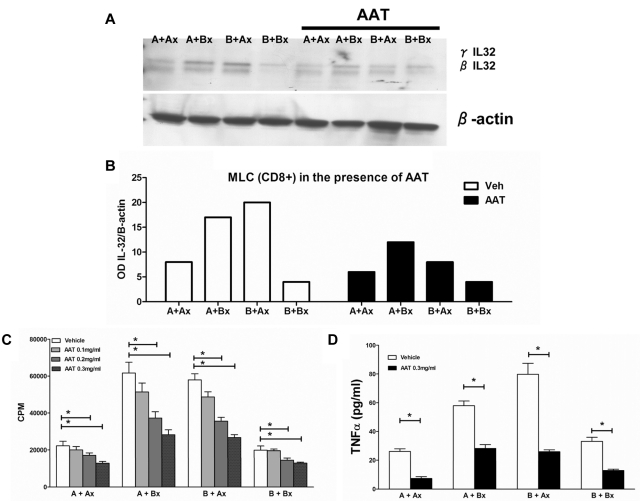
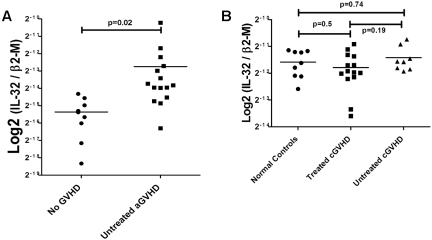



References
-
- Antin JH, Ferrara JLM. Cytokine dysregulation and acute graft-versus-host disease. Blood. 1992;80(12):2964–2968. - PubMed
-
- Stirewalt DL, Mhyre AJ, Marcondes M, et al. Tumour necrosis factor-induced gene expression in human marrow stroma: clues to the pathophysiology of MDS? Br J Haematol. 2008;140(4):444–453. - PubMed
-
- Hong J, Bae S, Kang Y, et al. Suppressing IL-32 in monocytes impairs the induction of the proinflammatory cytokines TNFalpha and IL-1beta. Cytokine. 2010;49(2):171–176. - PubMed
-
- Mun SH, Kim JW, Nah SS, et al. Tumor necrosis factor alpha-induced interleukin-32 is positively regulated via the Syk/protein kinase Cdelta/JNK pathway in rheumatoid synovial fibroblasts. Arthritis Rheum. 2009;60(3):678–685. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL094260/HL/NHLBI NIH HHS/United States
- HL094260/HL/NHLBI NIH HHS/United States
- HL095999/HL/NHLBI NIH HHS/United States
- P01 AI033484/AI/NIAID NIH HHS/United States
- K08 DK085156/DK/NIDDK NIH HHS/United States
- R56 AI015614/AI/NIAID NIH HHS/United States
- K08 DK085156-01/DK/NIDDK NIH HHS/United States
- P01 HL036444/HL/NHLBI NIH HHS/United States
- R01 HL095999/HL/NHLBI NIH HHS/United States
- AI-15614/AI/NIAID NIH HHS/United States
- HL036444/HL/NHLBI NIH HHS/United States
- AI33484/AI/NIAID NIH HHS/United States
- R01 AI015614/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
