

Upregulation of KCC2 activity by zinc-mediated neurotransmission via the mZnR/GPR39 receptor
- PMID: 21900570
- PMCID: PMC3227684
- DOI: 10.1523/JNEUROSCI.2205-11.2011
Upregulation of KCC2 activity by zinc-mediated neurotransmission via the mZnR/GPR39 receptor
Abstract
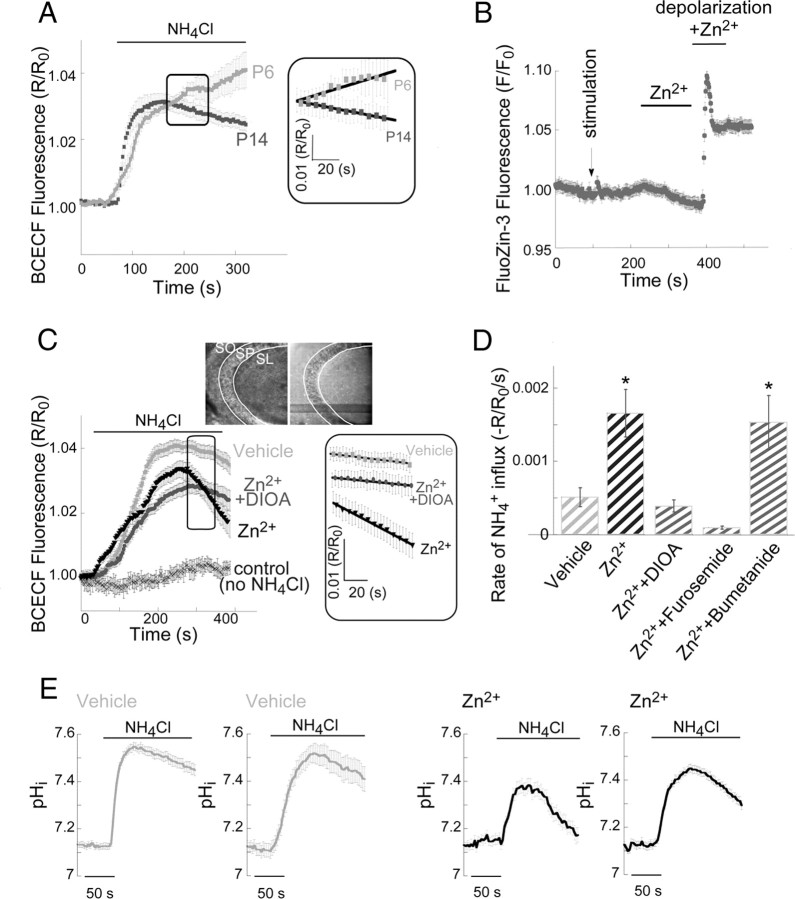
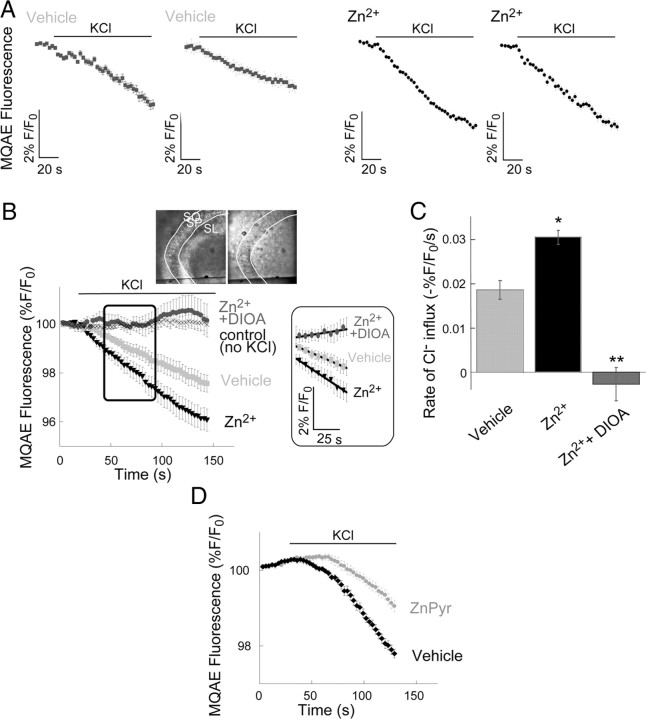
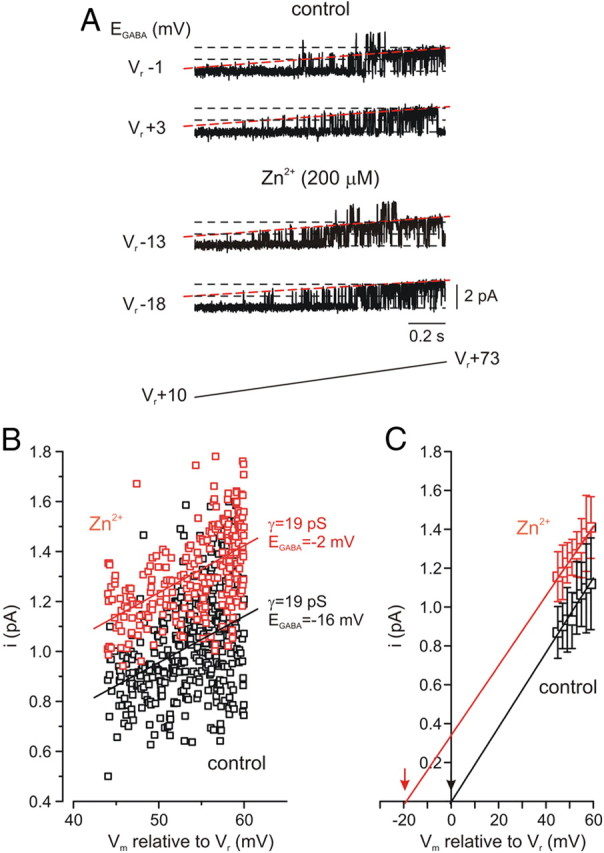
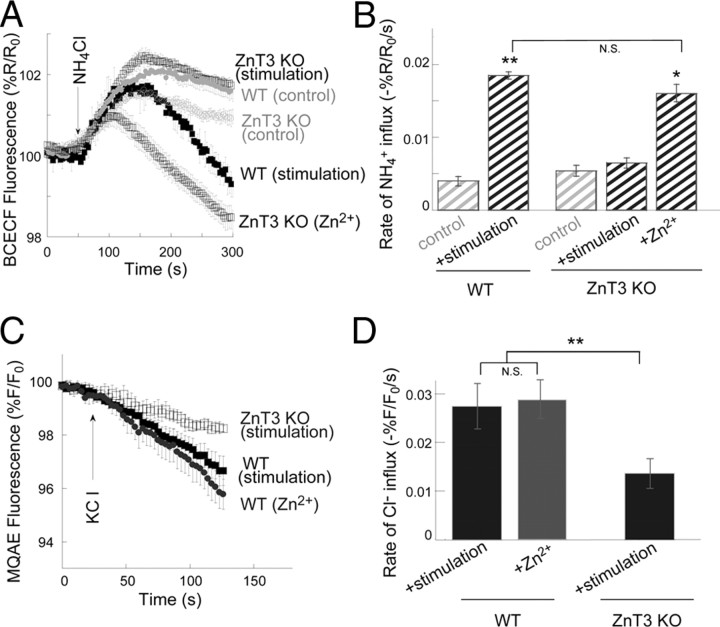
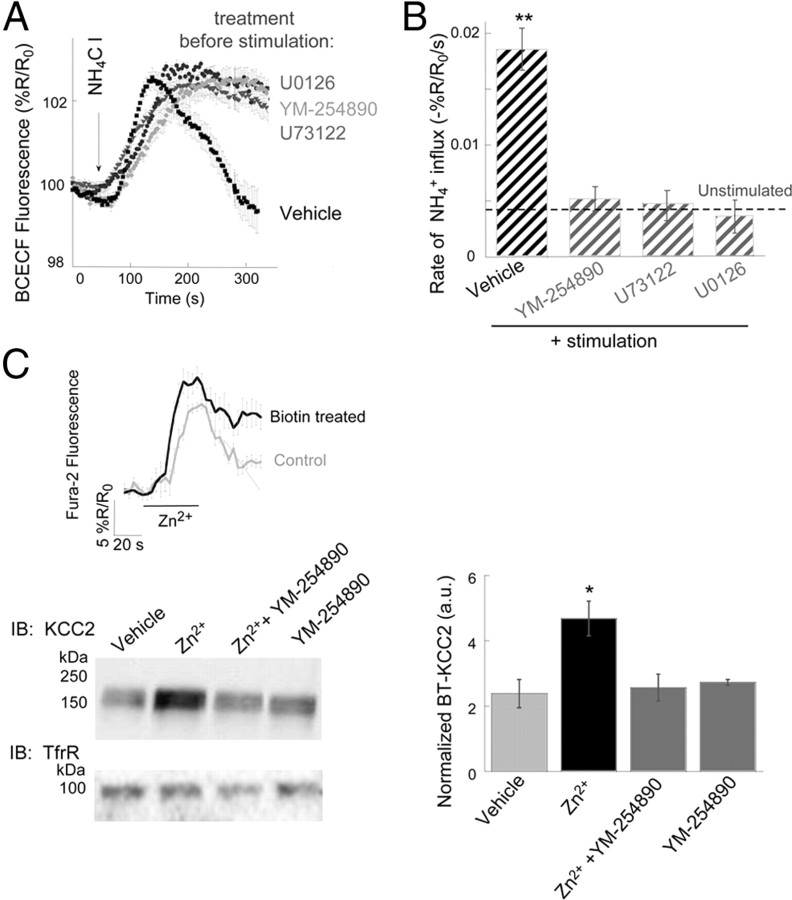
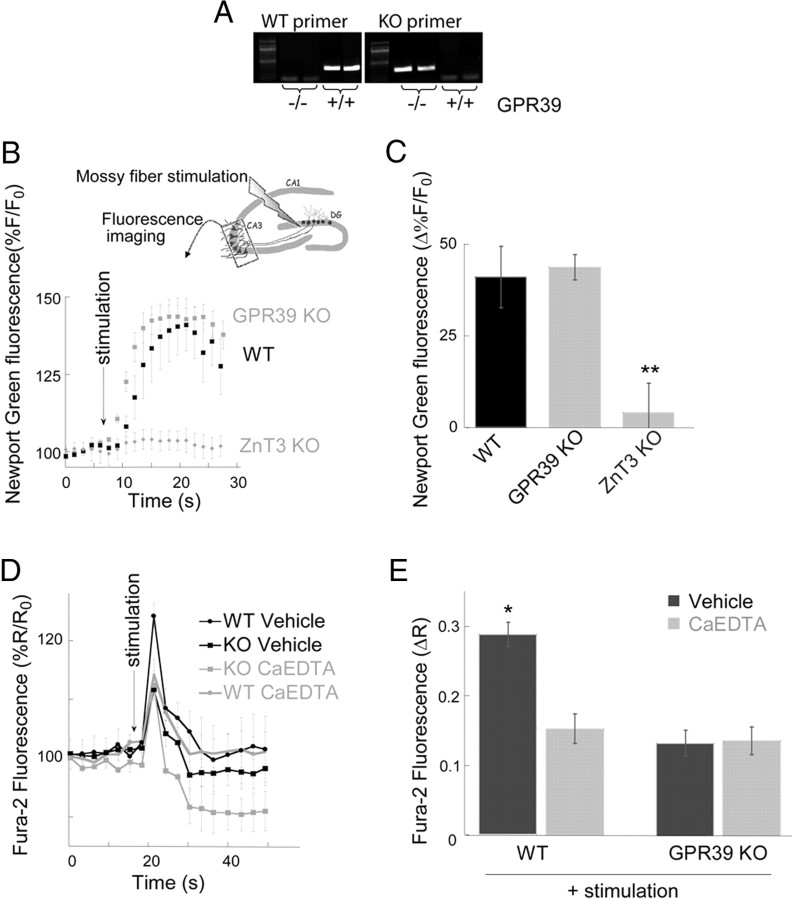
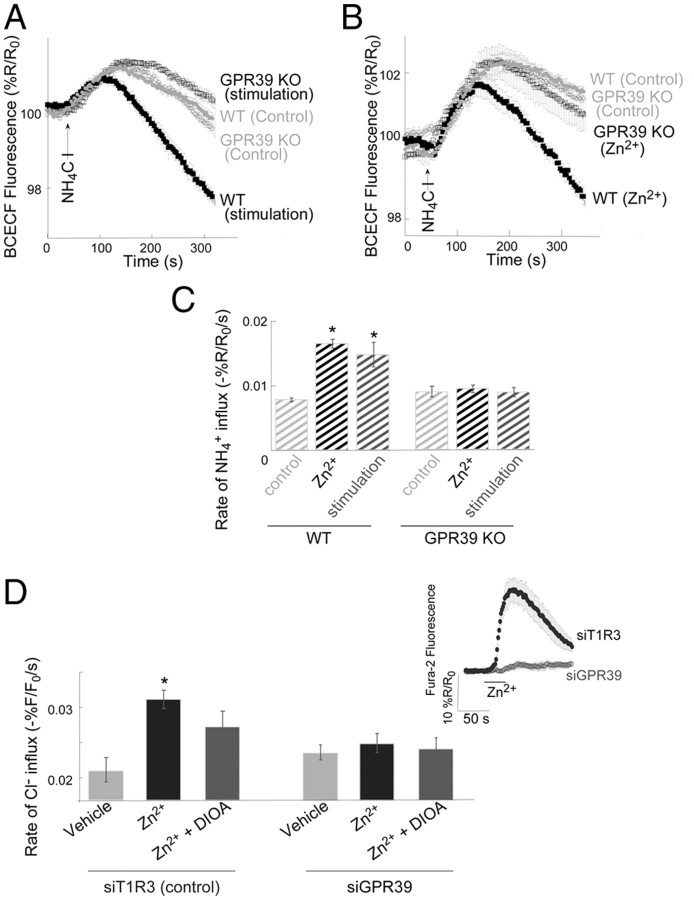
Vesicular Zn(2+) regulates postsynaptic neuronal excitability upon its corelease with glutamate. We previously demonstrated that synaptic Zn(2+) acts via a distinct metabotropic zinc-sensing receptor (mZnR) in neurons to trigger Ca(2+) responses in the hippocampus. Here, we show that physiological activation of mZnR signaling induces enhanced K(+)/Cl(-) cotransporter 2 (KCC2) activity and surface expression. As KCC2 is the major Cl(-) outward transporter in neurons, Zn(2+) also triggers a pronounced hyperpolarizing shift in the GABA(A) reversal potential. Mossy fiber stimulation-dependent upregulation of KCC2 activity is eliminated in slices from Zn(2+) transporter 3-deficient animals, which lack synaptic Zn(2+). Importantly, activity-dependent ZnR signaling and subsequent enhancement of KCC2 activity are also absent in slices from mice lacking the G-protein-coupled receptor GPR39, identifying this protein as the functional neuronal mZnR. Our work elucidates a fundamentally important role for synaptically released Zn(2+) acting as a neurotransmitter signal via activation of a mZnR to increase Cl(-) transport, thereby enhancing inhibitory tone in postsynaptic cells.
Figures
References
-
- Aizenman E, Stout AK, Hartnett KA, Dineley KE, McLaughlin B, Reynolds IJ. Induction of neuronal apoptosis by thiol oxidation: putative role of intracellular zinc release. J Neurochem. 2000;75:1878–1888. - PubMed
-
- Amaral DG, Witter MP. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience. 1989;31:571–591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous