

Lysophosphatidic Acid Stimulates the Proliferation of Ovarian Cancer Cells via the gep Proto-Oncogene Gα(12)
- PMID: 21901169
- PMCID: PMC3161422
- DOI: 10.1177/1947601911419362
Lysophosphatidic Acid Stimulates the Proliferation of Ovarian Cancer Cells via the gep Proto-Oncogene Gα(12)
Abstract
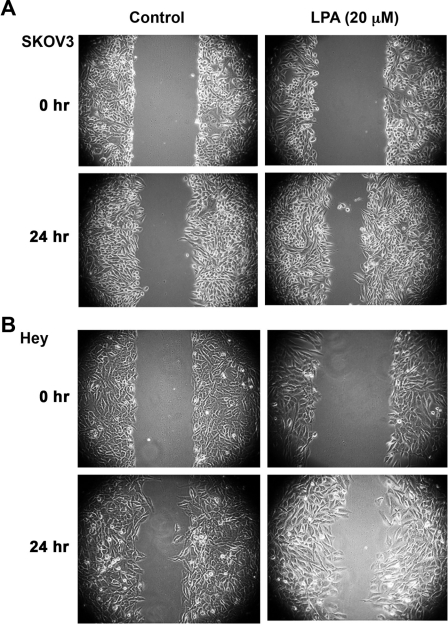
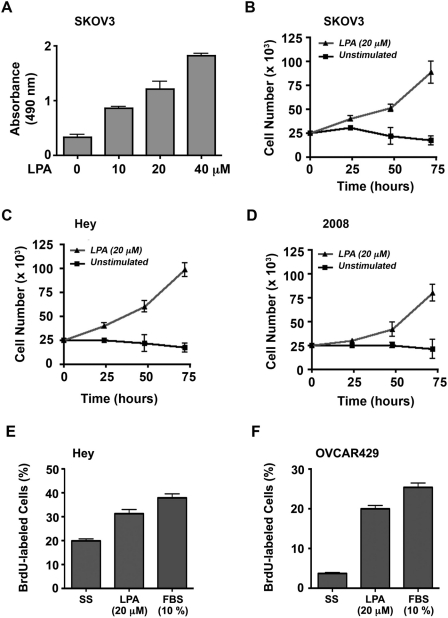
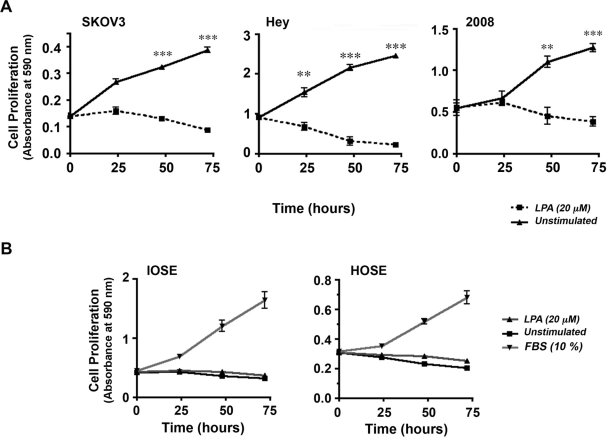
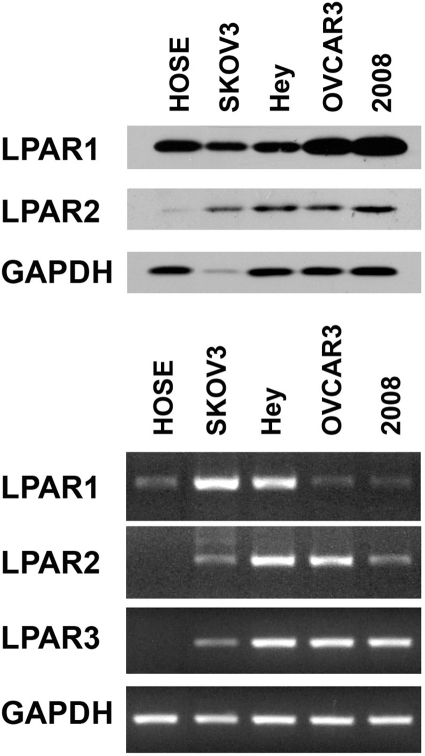
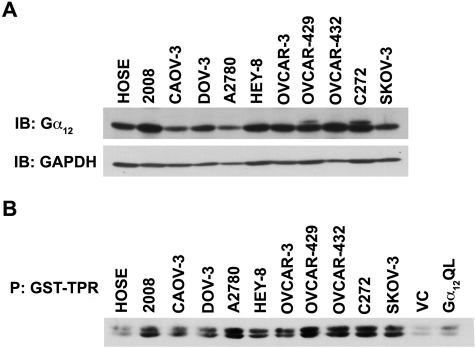
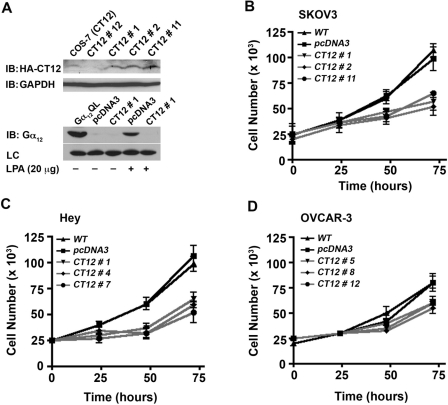
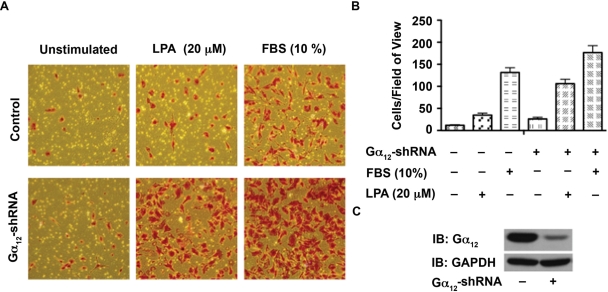
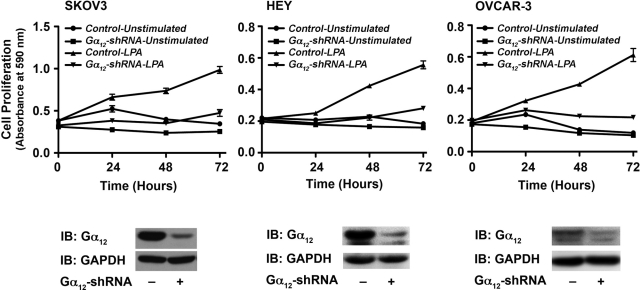
Lysophosphatidic acid (LPA), an agonist that activates specific G protein-coupled receptors, is present at an elevated concentration in the serum and ascitic fluid of ovarian cancer patients. Although the increased levels of LPA have been linked to the genesis and progression of different cancers including ovarian carcinomas, the specific signaling conduit utilized by LPA in promoting different aspects of oncogenic growth has not been identified. Here, we show that LPA stimulates both migration and proliferation of ovarian cancer cells. Using multiple approaches, we demonstrate that the stimulation of ovarian cancer cells with LPA results in a robust and statistically significant proliferative response. Our results also indicate that Gα(12), the gep proto-oncogene, which can be stimulated by LPA via specific LPA receptors, is overtly activated in a large array of ovarian cancer cells. We further establish that LPA stimulates the rapid activation of Gα(12) in SKOV-3 cells and the expression of CT12, an inhibitory minigene of Gα(12) that disrupts LPAR-Gα(12) interaction and potently inhibits such activation. Using this inhibitory molecule as well as the shRNA approach, we show that the inhibition of Gα(12) or silencing of its expression drastically and significantly attenuates LPA-mediated proliferation of ovarian cancer cell lines such as SKOV3, Hey, and OVCAR-3. Together with our findings that the silencing of Gα(12) does not have any significant effect on LPA-mediated migratory response of SKOV3 cells, our results point to a critical role for LPA-LPAR-Gα(12) signaling in ovarian cancer cell proliferation and not in migration. Thus, results presented here for the first time demonstrate that the gep proto-oncogene forms a specific node in LPA-LPAR-mediated mitogenic signaling in ovarian cancer cells.
Keywords: LPA; gep; metastasis; oncogene; proliferation.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
Similar articles
-
The gep proto-oncogene Gα12 mediates LPA-stimulated activation of CREB in ovarian cancer cells.Cell Signal. 2014 Jan;26(1):122-32. doi: 10.1016/j.cellsig.2013.08.012. Epub 2013 Sep 19. Cell Signal. 2014. PMID: 24055910 Free PMC article.
-
The gep proto-oncogene Gα13 mediates lysophosphatidic acid-mediated migration of pancreatic cancer cells.Pancreas. 2013 Jul;42(5):819-28. doi: 10.1097/MPA.0b013e318279c577. Pancreas. 2013. PMID: 23508014 Free PMC article.
-
Endogenous RGS proteins attenuate Galpha(i)-mediated lysophosphatidic acid signaling pathways in ovarian cancer cells.Cell Signal. 2008 Feb;20(2):381-9. doi: 10.1016/j.cellsig.2007.10.026. Epub 2007 Nov 7. Cell Signal. 2008. PMID: 18083345
-
Lysophosphatidic acid (LPA) signalling in cell migration and cancer invasion: a focussed review and analysis of LPA receptor gene expression on the basis of more than 1700 cancer microarrays.Biol Cell. 2013 Aug;105(8):317-33. doi: 10.1111/boc.201300011. Epub 2013 Jun 3. Biol Cell. 2013. PMID: 23611148 Review.
-
Beta-catenin, cancer, and G proteins: not just for frizzleds anymore.Sci STKE. 2005 Jul 12;2005(292):pe35. doi: 10.1126/stke.2922005pe35. Sci STKE. 2005. PMID: 16014605 Review.
Cited by
-
Synthesis and Biological Evaluation of Lysophosphatidic Acid Analogues Using Conformational Restriction and Bioisosteric Replacement Strategies.ACS Omega. 2023 Dec 11;8(51):49278-49288. doi: 10.1021/acsomega.3c07668. eCollection 2023 Dec 26. ACS Omega. 2023. PMID: 38162765 Free PMC article.
-
Tumor necrosis factor receptor modulator spermatogenesis-associated protein 2 is a novel predictor of outcome in ovarian cancer.Cancer Sci. 2019 Mar;110(3):1117-1126. doi: 10.1111/cas.13955. Epub 2019 Feb 16. Cancer Sci. 2019. PMID: 30697874 Free PMC article.
-
LPA signaling is regulated through the primary cilium: a novel target in glioblastoma.Oncogene. 2018 Mar;37(11):1457-1471. doi: 10.1038/s41388-017-0049-3. Epub 2018 Jan 11. Oncogene. 2018. PMID: 29321663 Free PMC article.
-
Hax-1 is required for Rac1-Cortactin interaction and ovarian carcinoma cell migration.Genes Cancer. 2014 Mar;5(3-4):84-99. doi: 10.18632/genesandcancer.8. Genes Cancer. 2014. PMID: 25053987 Free PMC article.
-
Lysophosphatidic acid activates lipogenic pathways and de novo lipid synthesis in ovarian cancer cells.J Biol Chem. 2012 Jul 20;287(30):24990-5000. doi: 10.1074/jbc.M112.340083. Epub 2012 Jun 3. J Biol Chem. 2012. PMID: 22665482 Free PMC article.
References
-
- Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60:277-300 - PubMed
-
- Mills GB, May C, McGill M, Roifman CM, Mellors A. A putative new growth factor in ascitic fluid from ovarian cancer patients: identification, characterization, and mechanism of action. Cancer Res. 1998;48:1066-71 - PubMed
-
- Fishman DA, Liu Y, Ellerbroek SM, Stack MS. Lysophosphatidic acid promotes matrix metalloproteinase (MMP) activation and MMP-dependent invasion in ovarian cancer cells. Cancer Res. 2001;61:3194-9 - PubMed
-
- Sawada K, Morishige K, Tahara M, et al. Alendronate inhibits lysophosphatidic acid-induced migration of human ovarian cancer cells by attenuating the activation of rho. Cancer Res. 2002;62:6015-20 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous