

Review
doi: 10.1021/cr200021m.
Epub 2011 Sep 8.
Biosynthesis of cholesterol and other sterols
Affiliations
- PMID: 21902244
- PMCID: PMC3191736
- DOI: 10.1021/cr200021m
Item in Clipboard
Review
Biosynthesis of cholesterol and other sterols
Chem Rev.
.
Free PMC article
No abstract available
Figures

Common tetracyclic steroid frame containing the 1,2-cyclopentanoperhydrophenanthrene ring skeleton.

Perspective drawings of the cholesterol molecule showing four domains of functional importance (left) and the flat elongated structure presumed to form in the membrane (right).

Two systems recognized for numbering of carbon atoms of the sterol nucleus and side chain. The conventional system, which incorporates the α/β-chiral descriptors in nucleus and side chain is based on the Fieser and Fieser book Steroids and biosynthetic considerations as discussed in refs (, , and 5) (panel A), versus the 1989 IUPAC recommendations (panel B).

The cyclization of squalene-2,3-oxide (i) folded in either the chair–chair–chair–boat–unfolded (ii) or chair–boat–chair–boat–unfolded (ii) conformations to yield a cation (ii), which can stabilize to produce the dammarane, tirucallane, euphane, and cucurbitane skeletons (via path a) or cycloartane and lanostane skeletons (via path b). Generally, the cyclization products contain a 3β-OH group and a Δ8-bond or in the case of cucurbitacin a Δ5-bond.

The prenyl transferase reaction and formation of farnesyl diphosphate (FPP). Prenyl (C5) units composed of isopentyl diphosphates (IPP) are assembled in a head to tail fusion to yield C10 (geranyl diphosphate, GPP) and in repeat fashion to C15 (FPP) isoprenoids. Alternate isoprene unit assemblies yield a variety of different structures notable in terpene metabolism. Enz = enzyme and M++ can be Mn2+ or Mg2+ ions.

Proposed mechanism for the coupling of farnesyl diphosphate (FPP) to form squalene.

Distribution of acetate carbon atoms found in cholesterol; a repeating pattern of five carbon atoms (isoprene unit), surrounded by dotted lines, is recognizable in three places in the molecule.

Interpretation of the mechanism of squalene-2,3-oxide cyclization to lanosterol and cycloartenol according to refs (, , and 64). The cyclization mechanisms are hypothesized to require “X– group” (any electron-donating group on the enzyme) participation to generate the C20R configuration, which requires the side chain of the X– group bound protosteroid to rotate from left to right about C20 and formation of “bent” cycloartenol from a “flat” 9β,19-cyclosteroid intermediate. See text.

Overview of compartmentalized isoprenoid–sterol biosynthesis pathways. Feeding studies of [2-13C]leucine or [1-13C]glucose distinguish the leucine–HMGCoA, acetate–mevalonic acid (MVA) or the 2-C-methyl-d -erythritol-4-phosphate (MEP) pathways that contribute carbon to isopentenyl diphosphate (IPP). The 13C-labeling pattern of ergosterol (synthesized in all major eukaryote Kingdoms) reveals which of the pathways operate in a given organism and whether cross-talk between the pathways exist. In the forward direction, IPP converts to ergosterol or IPP can retroconvert via the MVA shunt to HMGCoA and to acetate. See text.

The cyclization of squalene-2,3-oxide (1) catalyzed by the lanosterol synthase (LAS) or cycloartenol synthase (CAS) to give true sterols. Ring annulation proceeds by synthase (cyclase)-specific cationic mechanisms involving a common protosteroid cation (17α-H/17β-side chain orientation, 3). The cyclization cascade terminates with formation of either (i) intermediate 4 (LAS) followed by its deprotonation at C9 or C7 to form lanosterol 11 or lanosta-7,24-dienol 12, respectively or (ii) intermediate 6 (CAS), which depending on the position of deprotonation at C7, C19, or C11 yields lanosta-7,24-dienol 13, cycloartenol 14, or parkeol 15, respectively; trans-cyclization of the chair–boat type of system in 6a (natural intermediate compared with 6b as discussed in the text) affords the 9β-H instead of a 9α-H, which is necessary to complete biosynthesis of cholesterol. The asymmetric conformation of bound intermediates 4 and 6 with a 20α-H-atom (20R-configuration) eclipsed to 17α-H-atom produce an enzyme-bound “left-handed” rotamer at C20. After release of true sterols 7–10 from the active site, the sterol side chain can rotate about C20, such that C21 and C22 lies in a 1,3-diaxial relationship with C18 to form the energetically favorable “right-handed” conformation of 11 to 15 utilized in membranes.

Structure of “flat” cycloartenol established by NMR (left) and X-ray (right) measurements.

A generalized route from cycloartenol 1 to Δ5-24α/β-ethyl sterols, stigmasterol 18 and poriferasterol 23, respectively.

The sterol alkylation reaction pathways operating in different organisms; routes 1 in fungi, protozoa, and plants to form C9 and C10 side chains and routes 2 and 3 to form C11 extended side chains in marine organisms.

Mechanisms for methyl transfer to Δ24(25)-substrate in the action of sterol C24-methyltransferase; dot shows to 13C-labeled carbon. In mechanisms A and C, the carbocationic intermediates are discrete species, and an important difference between the two mechanisms involves the SN2 (Si, β)-attack of methyl cation on the substrate double bond coupled to the regiospecific deprotonation at C28 to yield the 25S stereochemistry typical of 24-alkyl sterols of the ergostane and stigmastane family. In mechanism B, the formation of carbocation intermediates is avoided by postulating the participation of an enzymatic nucelophile-X to form covalent bonds with incipient cations to yield the C25R stereochemistry typical of animal cholesterol.

C1-Transfer mechanisms in the conversion of Δ24(28)-substrate into 24-ethyl(idene) products. In each case, the isotopically labeled substrate contains a 13C-27 atom (dot) and a Z- or E-2H-atom selectively introduced at C28. The “cis-process” has been eliminated by experiment.

Revised mechanism for the reduction of the sterol Δ24-double bond by the action of sterol C24-reductase (24-SR), which uses NADPH as coenzyme. The new hydrogen atoms (as hydrogen or tritium) introduced are marked with circles, and 13C-labeled carbon is shown with a dot. The mechanism requires introduction of a proton from an active site donor, for example, lysine, to the re-face of the substrate double bond at C24 (1) followed by trans-addition of the hydride ion to eliminate the carbonium ion at the si-face of C25 (2) thereby forming the C26 Pro-R stereochemistry typical of animal cholesterol (3).
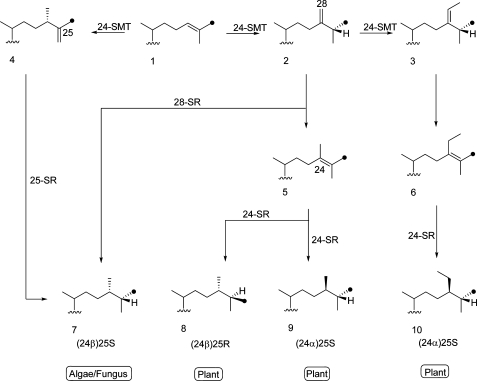
Phyla-specific pathways for the regio- and stereospecific reduction of a remote double bond in 24-alkyl sterol side chains. Three distinct sterol reductases are considered in the formation of the C24-methyl/ethyl and C25-H stereochemistry, sterol C28-reductase (28-SR), sterol C25-reductase (25-SR), and sterol C24-reductase (24-SR). Products 8, 9, and 10 can originate by the action of the same plant 24-SR. Dot indicates 13C27-labeled atom. See text.

Stereochemical scheme for the stepwise enzymatic removal of C4α-methyl group and reduction of C3 ketone to produce a 3β-hydroxyl group. The first set of enzymatic reactions (1–5) is catalyzed by C4-sterol methyl oxidase (4-SMO) in complex with O2, NADPH, and Cytb5 yielding a C4-oxygenated intermediate. The second set of reaction steps (5–7) involves C4-sterol decarboxylase (4-SDC) [3β-hydroxysteroid dehydrogenase/C4-decarboxylase (4-HSD/D)] and NAD+ catalyzing sterol 3-ketone formation; 4-SMO and 4-SDC are coupled but separate reactions. The third enzymatic reaction (7–8) involves sterol 3-reductase (3-SR) in complex with NADPH yielding 3β-hydroxy sterol. In the conversion of a 4,4-dimethyl sterol (e.g., lanosterol) to a C4-mono methyl sterol (e.g., 31-norlanosterol), the C4β-methyl group in the substrate 1 epimerizes to become the C4α-methyl group 8 in the product as shown. See text.

Possible pathways for oxidative removal of the 14α-methyl group in sterol biosynthesis (R = sterol side chain; Hw+ is proton from water) according to Akhtar, Galylor, and cowokers.− The reaction is carried out by 14α-sterol demethylase (14-SDM, CYP51) and proceeds by oxidation of the 14α-methyl group to alcohol 2 and thence to the C32-aldehyde 4. During cholesterol biosynthesis in rat liver the loss of C32 as formaldehyde is accompanied by loss of the 15α-hydrogen to generate a Δ8,14-sterol 8. Akhtar et al.(190) proposed two possible concepts for the elimination of the 32-aldehyde shown in paths a and b, with path b considered to act under physiological conditions.

Chemical steps in the reaction of sterol 14(15)-reductase (14-SR) in complex with NADPH to form a Δ8-sterol. Reduction of the Δ14-bond introduces the hydride from NADPH into the 14α-position and a proton from water (Hw+) into the 15β-position of 3. This results in an inversion of the 15β-H derived from the original 14α-methyl sterol precursor (Figure 19) into the 15α-position of the product Δ8-sterol.

Enzymatic processing of sterol 8(9)-bond to the 5-position in rings A and B of the sterol nucleus. The sequence considered is based on the catalytic competence of the sterol C8-to-C7 isomerase (8-SI), sterol-5-desaturase (5-SD), and sterol C7-reductase (7-SR) enzymes tested from animal, plant, and fungi. The stereospecificities of the hydrogen losses and additions in the reactions shown have been elucidated in rat liver and a few other organisms with regard to both the hydrogen of the sterol substrate and the NADPH coenzyme. Some variations not shown in the mechanism may exist. See text.

Cleavage of the 9β,19-cyclopropane ring system to form a Δ8-sterol by sterol 9β,19-cyclopropane mutase (19-SCM) according to Rahier and co-workers.(229) In order for the reaction to proceed in a 1,2-trans manner, it has been postulated that a negatively charged group (X–) associated with the cycloeucalenol 1 to obtusifoliol 3 isomerase (COI; 19-SCM) may participate as shown., The preferred substrate recognized by COI is a 4α-methyl 24(28)-methylene sterol, such as cycloeucalenol. The loss of a proton from C11 instead of 8β-H would produce a Δ9(11)-14α-methyl ring system, and this may also be a route to this type of sterol in plants and marine organisms.(12)

Two mechanisms for the cis-dehydrogenation reaction catalyzed by sterol C22-desaturase (22-SD), also known as CYP51A1 for the fungal enzyme and CYP107A for the plant enzyme, that leads to the introduction of the C22(23)-E-double bond in the sterol side chain; HR and HS refer to hydrogen atoms originally part of the MVA molecule.
References
-
- Myant N. B.The Biology of Cholesterol and Related Steroids; William Heinemann Medical Books Ltd.: London, 1981; p 1.
-
- Nes W. R.; McKean M. L.. Biochemistry of Steroids and Other Isopentenoids; University Park Press: Baltimore, MD, 1977; p 1.
-
- Brown M. S.; Goldstein. J. L. Science 1986, 232, 34. - PubMed
- http://nobelprize.org/.
-
- Gibbons G. F. Lipids 2002, 37, 1153. - PubMed
-
- Popjak G. In Lipids: Chemistry, Biochemistry and Nutrition; Mead J. F., Alfin-Slater R. B., Howton D. R., Popjak. G., Eds.; Plenum Press: New York, 1986; p 295.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
