

Chromosome organization by a nucleoid-associated protein in live bacteria
- PMID: 21903814
- PMCID: PMC3329943
- DOI: 10.1126/science.1204697
Chromosome organization by a nucleoid-associated protein in live bacteria
Abstract
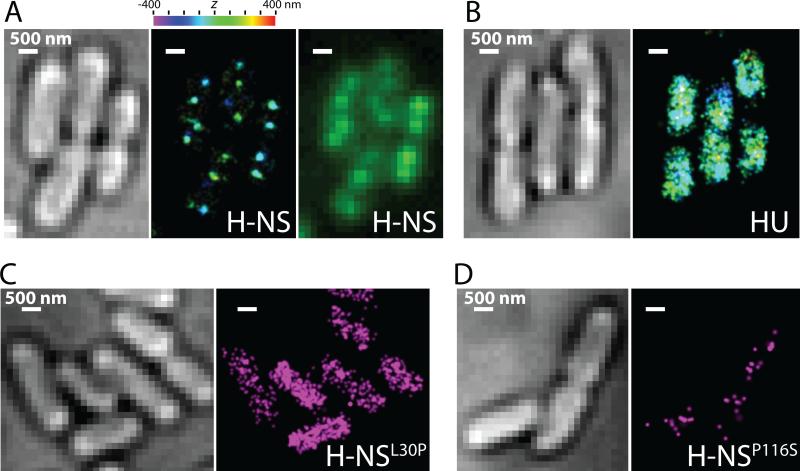
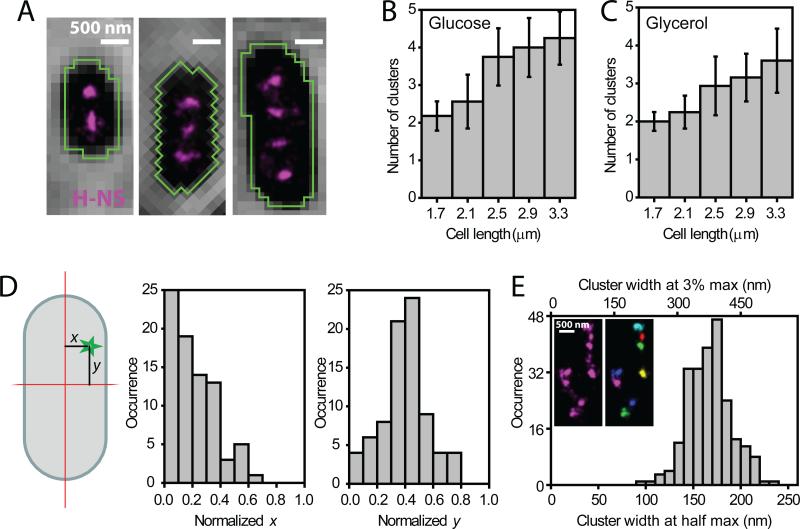
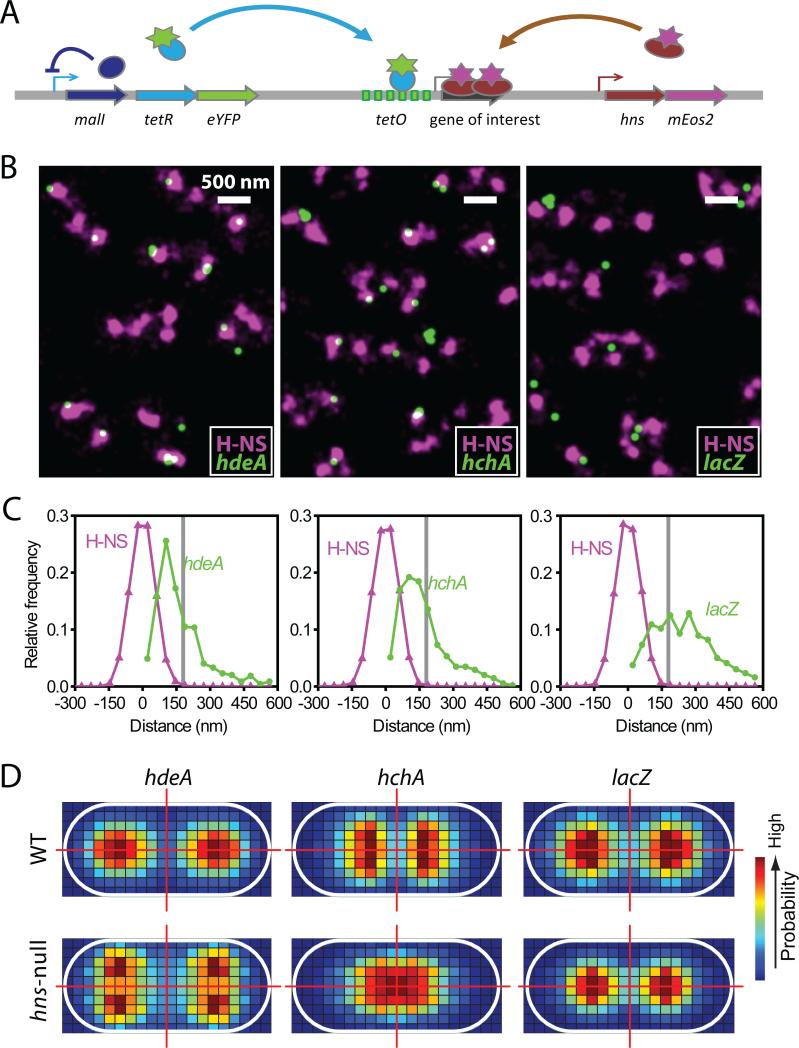
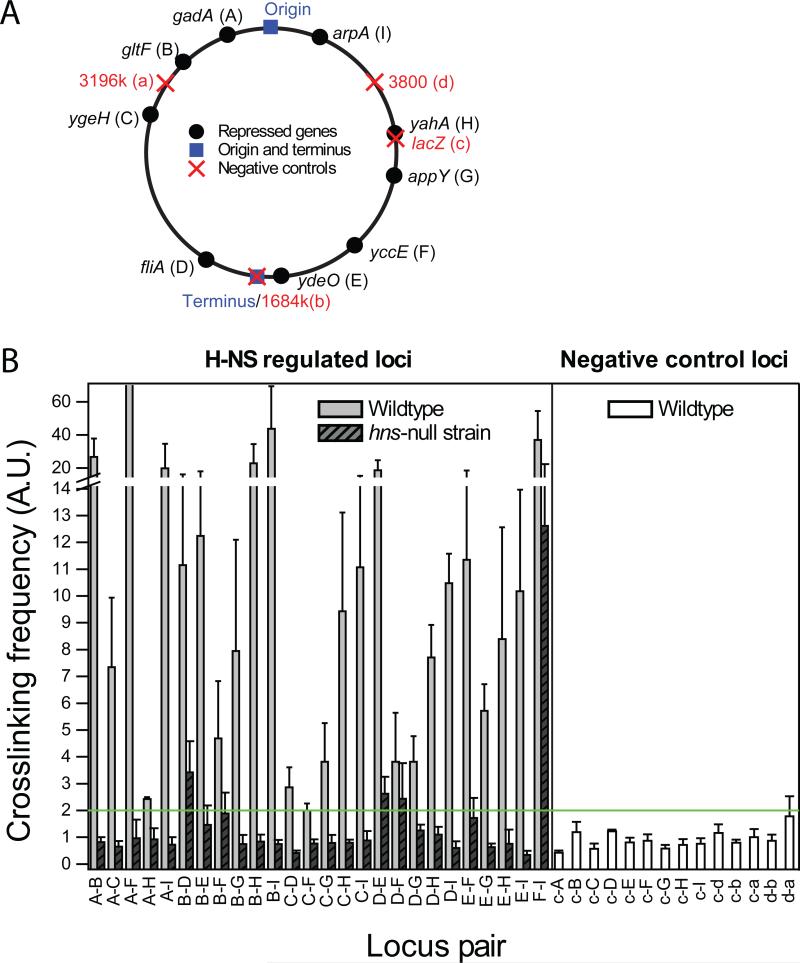
Bacterial chromosomes are confined in submicrometer-sized nucleoids. Chromosome organization is facilitated by nucleoid-associated proteins (NAPs), but the mechanisms of action remain elusive. In this work, we used super-resolution fluorescence microscopy, in combination with a chromosome-conformation capture assay, to study the distributions of major NAPs in live Escherichia coli cells. Four NAPs--HU, Fis, IHF, and StpA--were largely scattered throughout the nucleoid. In contrast, H-NS, a global transcriptional silencer, formed two compact clusters per chromosome, driven by oligomerization of DNA-bound H-NS through interactions mediated by the amino-terminal domain of the protein. H-NS sequestered the regulated operons into these clusters and juxtaposed numerous DNA segments broadly distributed throughout the chromosome. Deleting H-NS led to substantial chromosome reorganization. These observations demonstrate that H-NS plays a key role in global chromosome organization in bacteria.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
