

Homeostatic control of neural activity: a Drosophila model for drug tolerance and dependence
- PMID: 21906535
- PMCID: PMC4862361
- DOI: 10.1016/B978-0-12-387003-2.00002-1
Homeostatic control of neural activity: a Drosophila model for drug tolerance and dependence
Abstract
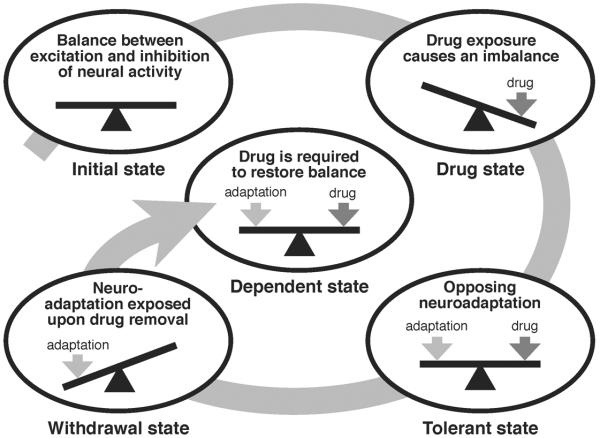
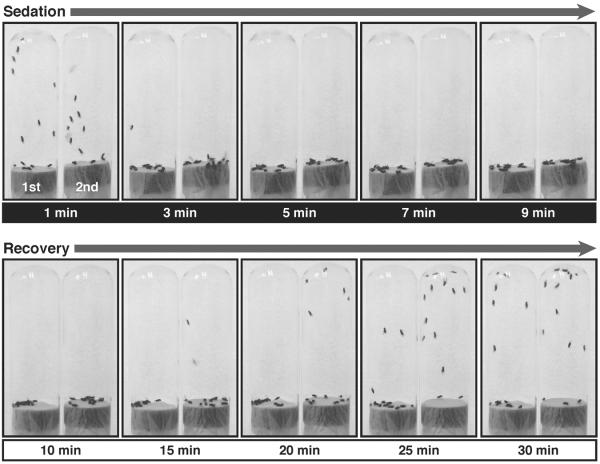
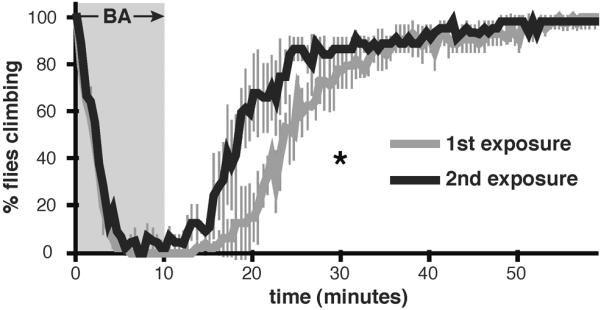
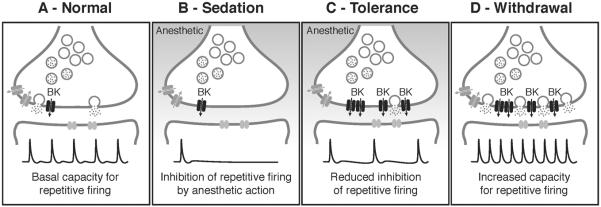
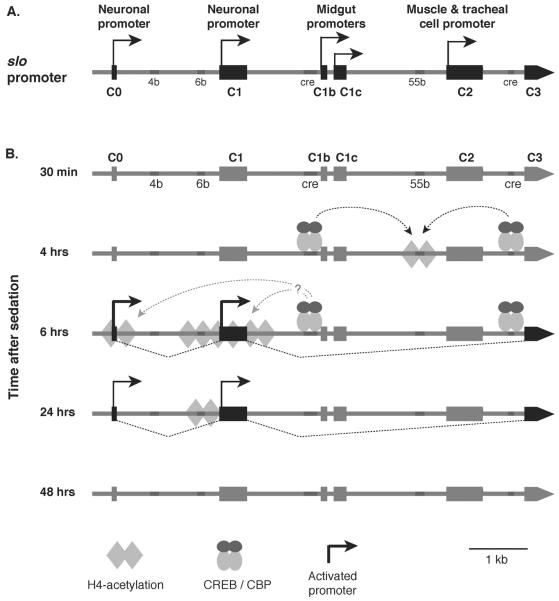
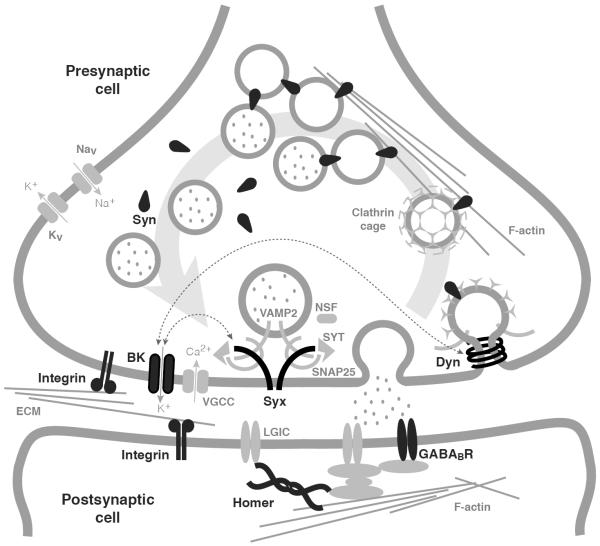
Drug addiction is a complex condition of compulsive drug use that results in devastating physical and social consequences. Drosophila melanogaster has recently emerged as a valuable genetic model for investigating the mechanisms of addiction. Drug tolerance is a measurable endophenotype of addiction that can be easily generated and detected in animal models. The counteradaptive theory for drug dependence postulates that the homeostatic adaptations that produce drug tolerance become counteradaptive after drug clearance, resulting in symptoms of dependence. In flies, a single sedation with ethanol or with an organic solvent anesthetic (benzyl alcohol) induces functional tolerance, an adaptation of the nervous system that reduces the effect of these neural depressants. Here we review the role of the BK channel gene (slo) and genes that encode other synaptic proteins in the process of producing functional tolerance. These proteins are predicted to be part of an orchestrated response that involves specific interactions across a highly complex synaptic protein network. The response of the slo gene to drug exposure and the consequence of induced slo expression fit nicely the tenets of the counteradaptive theory for drug tolerance and dependence. Induction of slo expression represents an adaptive process that generates tolerance because it enhances neuronal excitability, which counters the sedative effects of the drugs. After drug clearance, however, the increase in slo expression leads to an allostatic withdrawal state that is characterized by an increase in the susceptibility for seizure. Together, these results demonstrate a common origin for development of drug tolerance and withdrawal hyperexcitability in Drosophila.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Allada R, Nash HA. Drosophila melanogaster as a model for study of general anesthesia: the quantitative response to clinical anesthetics and alkanes. Anesth. Analg. 1993;77:19–26. - PubMed
-
- Allis CD, Jenuwein T, Reinberg D, Caparros M-L. Epigenetics. Cold Spring Harbor Laboratory Press; Woodbury, NY: 2007.
-
- Ankney PF. A note on barometric pressure and behavior in Drosophila pseudoobscura. Behav. Genet. 1984;14:315–317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
