

Acetylation of yeast AMPK controls intrinsic aging independently of caloric restriction
- PMID: 21906795
- PMCID: PMC3176974
- DOI: 10.1016/j.cell.2011.07.044
Acetylation of yeast AMPK controls intrinsic aging independently of caloric restriction
Abstract
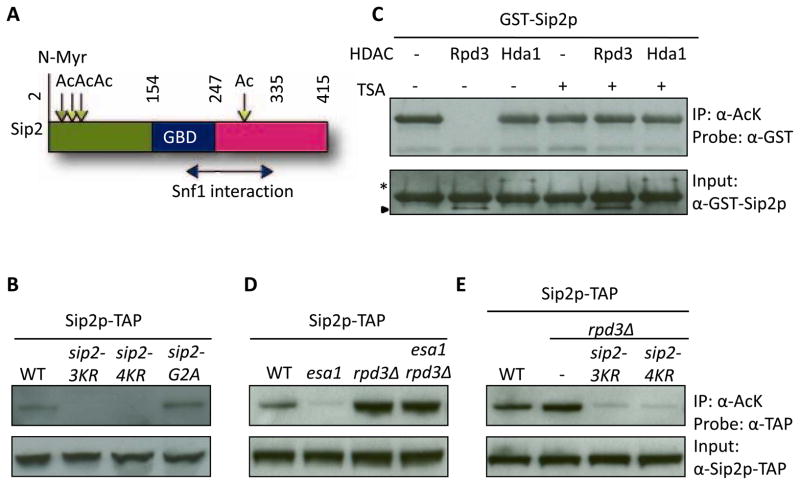
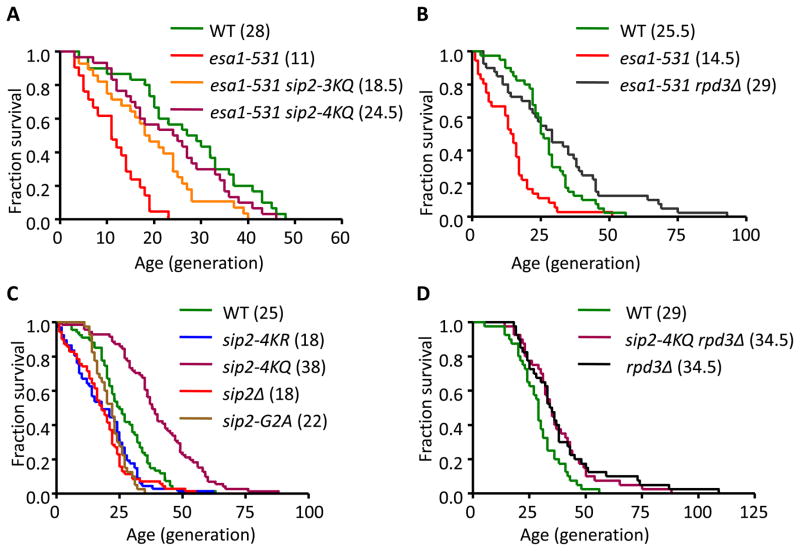
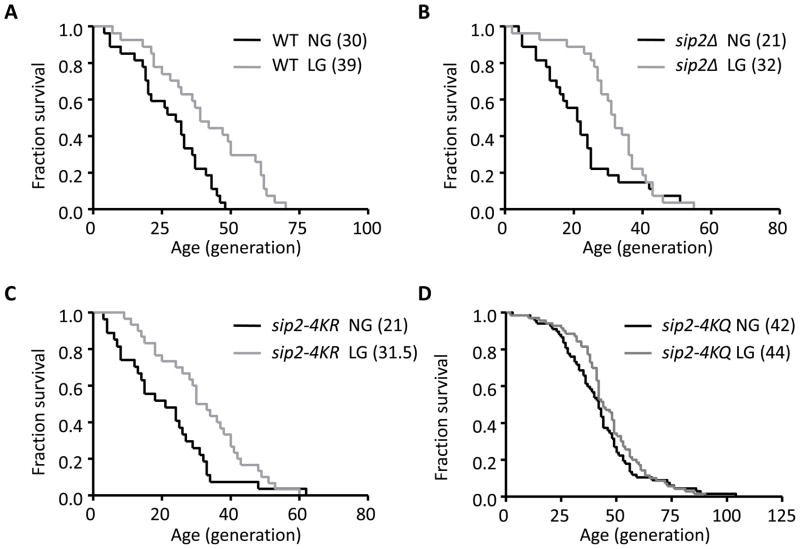
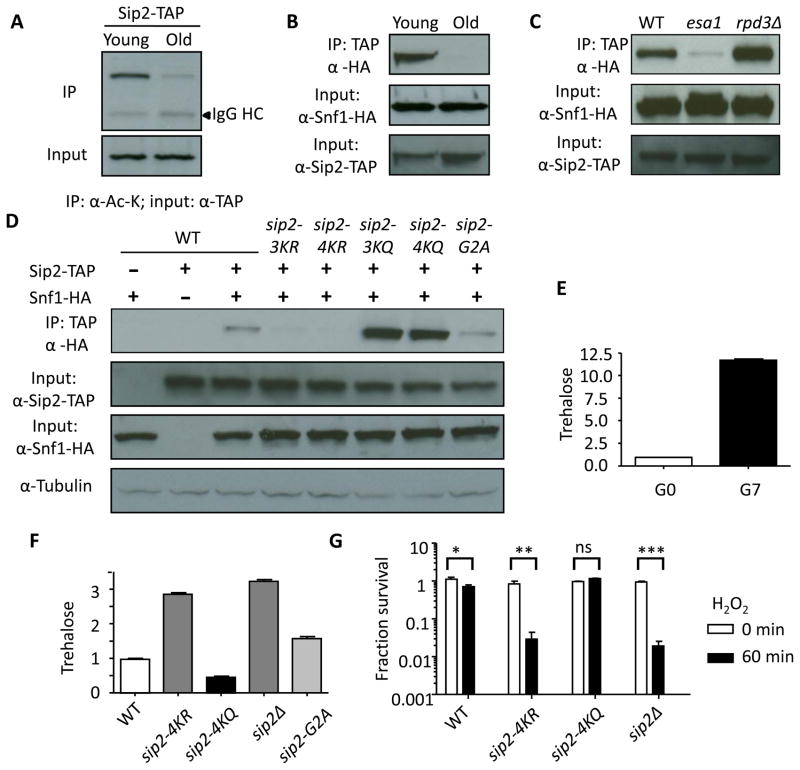
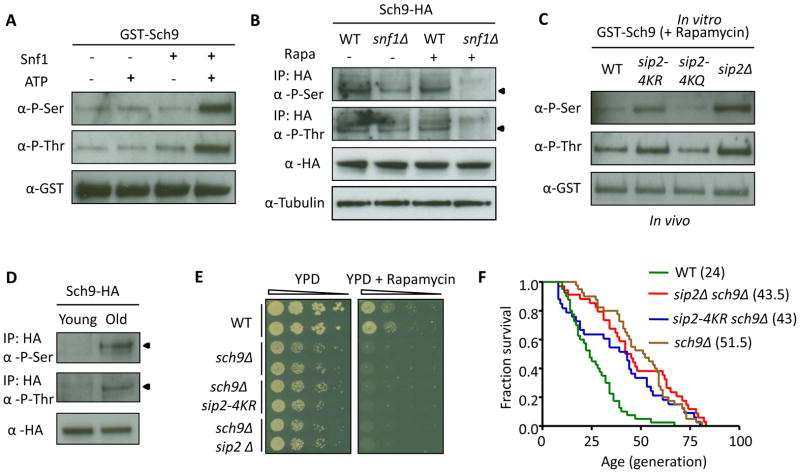
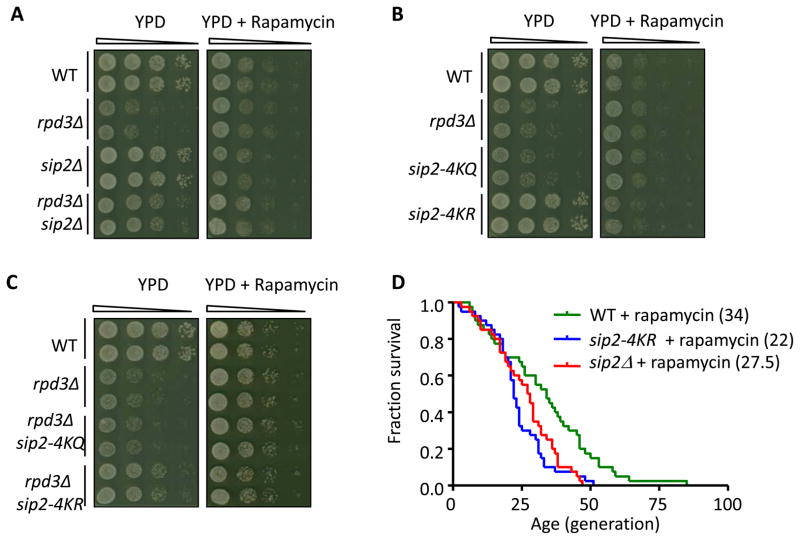
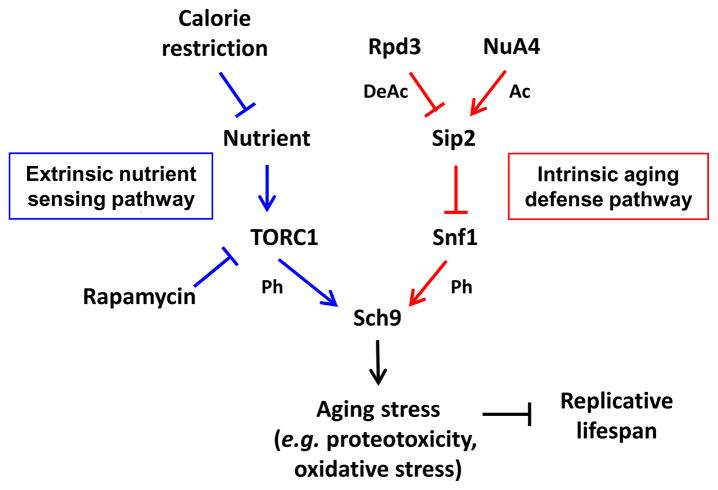
Acetylation of histone and nonhistone proteins is an important posttranslational modification affecting many cellular processes. Here, we report that NuA4 acetylation of Sip2, a regulatory β subunit of the Snf1 complex (yeast AMP-activated protein kinase), decreases as cells age. Sip2 acetylation, controlled by antagonizing NuA4 acetyltransferase and Rpd3 deacetylase, enhances interaction with Snf1, the catalytic subunit of Snf1 complex. Sip2-Snf1 interaction inhibits Snf1 activity, thus decreasing phosphorylation of a downstream target, Sch9 (homolog of Akt/S6K), and ultimately leading to slower growth but extended replicative life span. Sip2 acetylation mimetics are more resistant to oxidative stress. We further demonstrate that the anti-aging effect of Sip2 acetylation is independent of extrinsic nutrient availability and TORC1 activity. We propose a protein acetylation-phosphorylation cascade that regulates Sch9 activity, controls intrinsic aging, and extends replicative life span in yeast.
Copyright © 2011 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Comment in
-
SIP-ing the elixir of youth.Cell. 2011 Sep 16;146(6):859-60. doi: 10.1016/j.cell.2011.08.026. Cell. 2011. PMID: 21925309
Similar articles
-
Sip2, an N-myristoylated beta subunit of Snf1 kinase, regulates aging in Saccharomyces cerevisiae by affecting cellular histone kinase activity, recombination at rDNA loci, and silencing.J Biol Chem. 2003 Apr 11;278(15):13390-7. doi: 10.1074/jbc.M212818200. Epub 2003 Jan 31. J Biol Chem. 2003. PMID: 12562756
-
Ubp8 and SAGA regulate Snf1 AMP kinase activity.Mol Cell Biol. 2011 Aug;31(15):3126-35. doi: 10.1128/MCB.01350-10. Epub 2011 May 31. Mol Cell Biol. 2011. PMID: 21628526 Free PMC article.
-
The Snf1 protein kinase and its activating subunit, Snf4, interact with distinct domains of the Sip1/Sip2/Gal83 component in the kinase complex.Mol Cell Biol. 1997 Apr;17(4):2099-106. doi: 10.1128/MCB.17.4.2099. Mol Cell Biol. 1997. PMID: 9121458 Free PMC article.
-
AMPK in Yeast: The SNF1 (Sucrose Non-fermenting 1) Protein Kinase Complex.Exp Suppl. 2016;107:353-374. doi: 10.1007/978-3-319-43589-3_14. Exp Suppl. 2016. PMID: 27812987 Review.
-
AMPK/Snf1 signaling regulates histone acetylation: Impact on gene expression and epigenetic functions.Cell Signal. 2016 Aug;28(8):887-95. doi: 10.1016/j.cellsig.2016.03.009. Epub 2016 Mar 20. Cell Signal. 2016. PMID: 27010499 Review.
Cited by
-
Protein post-translational modifications in the regulation of cancer hallmarks.Cancer Gene Ther. 2023 Apr;30(4):529-547. doi: 10.1038/s41417-022-00464-3. Epub 2022 Apr 7. Cancer Gene Ther. 2023. PMID: 35393571 Review.
-
Mechanisms by which PE21, an extract from the white willow Salix alba, delays chronological aging in budding yeast.Oncotarget. 2019 Oct 8;10(56):5780-5816. doi: 10.18632/oncotarget.27209. eCollection 2019 Oct 8. Oncotarget. 2019. PMID: 31645900 Free PMC article.
-
Conventional and emerging roles of the energy sensor Snf1/AMPK in Saccharomyces cerevisiae.Microb Cell. 2018 Sep 29;5(11):482-494. doi: 10.15698/mic2018.11.655. Microb Cell. 2018. PMID: 30483520 Free PMC article. Review.
-
D-Xylose Sensing in Saccharomyces cerevisiae: Insights from D-Glucose Signaling and Native D-Xylose Utilizers.Int J Mol Sci. 2021 Nov 17;22(22):12410. doi: 10.3390/ijms222212410. Int J Mol Sci. 2021. PMID: 34830296 Free PMC article. Review.
-
Dietary natural products as epigenetic modifiers in aging-associated inflammation and disease.Nat Prod Rep. 2020 May 1;37(5):653-676. doi: 10.1039/c9np00057g. Epub 2020 Jan 29. Nat Prod Rep. 2020. PMID: 31993614 Free PMC article. Review.
References
-
- Amodeo GA, Rudolph MJ, Tong L. Crystal structure of the heterotrimer core of Saccharomyces cerevisiae AMPK homologue SNF1. Nature. 2007;449:492–495. - PubMed
-
- Barker MG, Brimage LJ, Smart KA. Effect of Cu, Zn superoxide dismutase disruption mutation on replicative senescence in Saccharomyces cerevisiae. FEMS Microbiol Lett. 1999;177:199–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
