

Pharmacological chaperones for misfolded gonadotropin-releasing hormone receptors
- PMID: 21907908
- PMCID: PMC3235646
- DOI: 10.1016/B978-0-12-385952-5.00008-7
Pharmacological chaperones for misfolded gonadotropin-releasing hormone receptors
Abstract
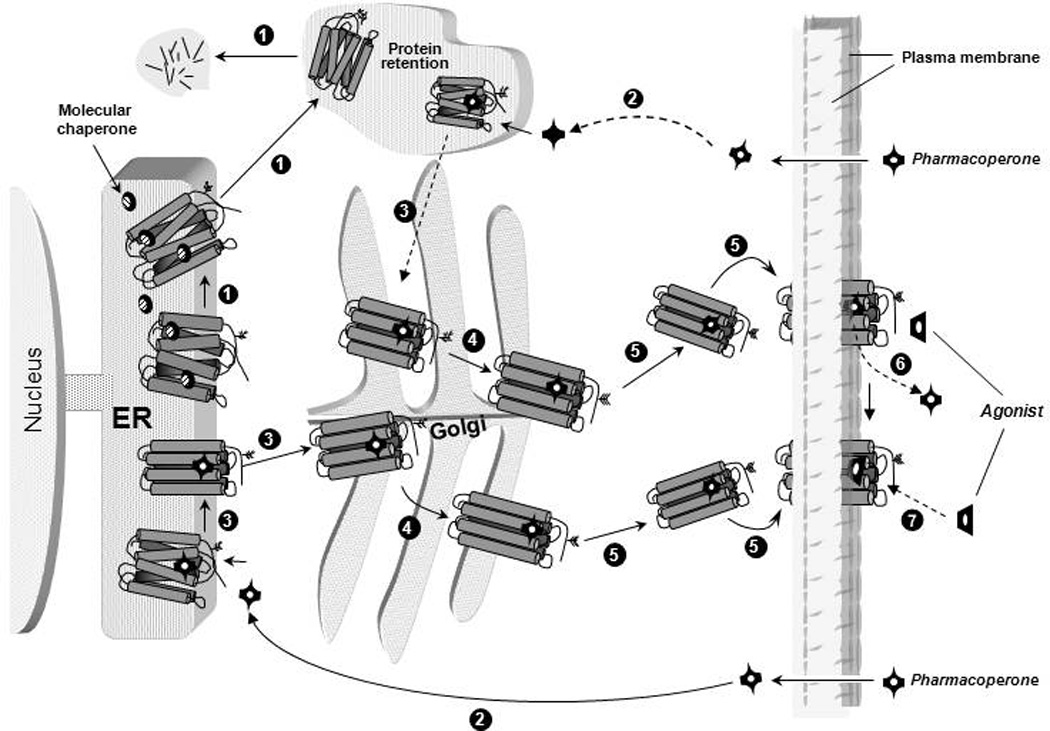
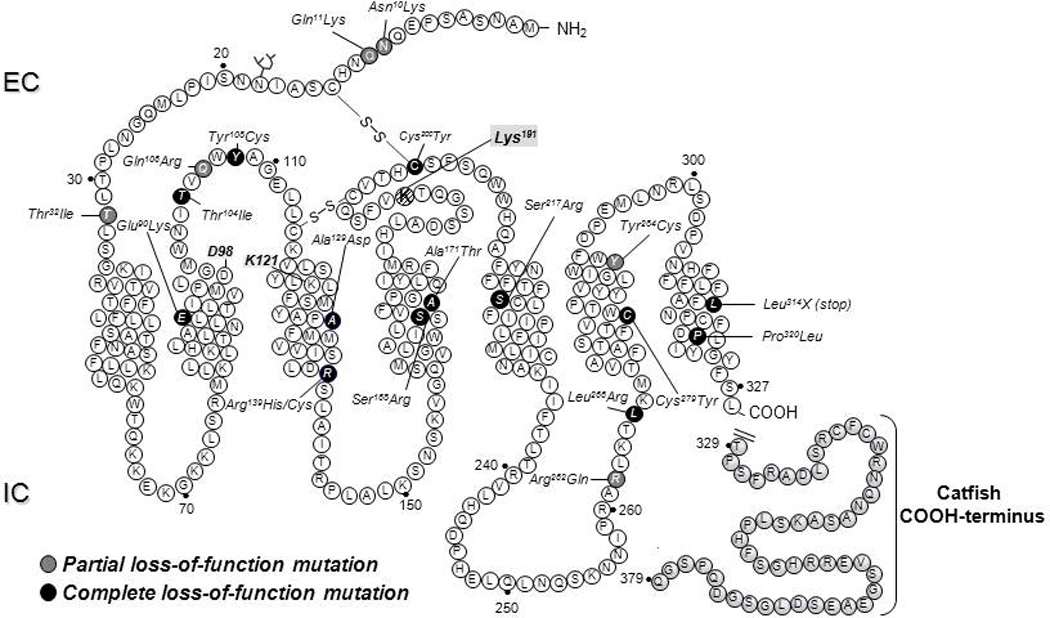
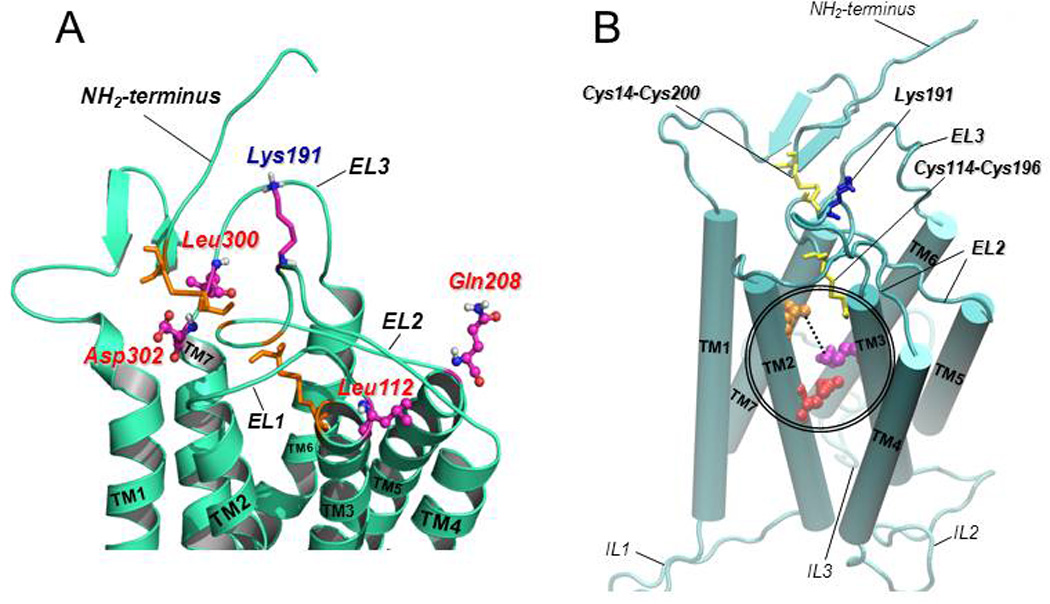
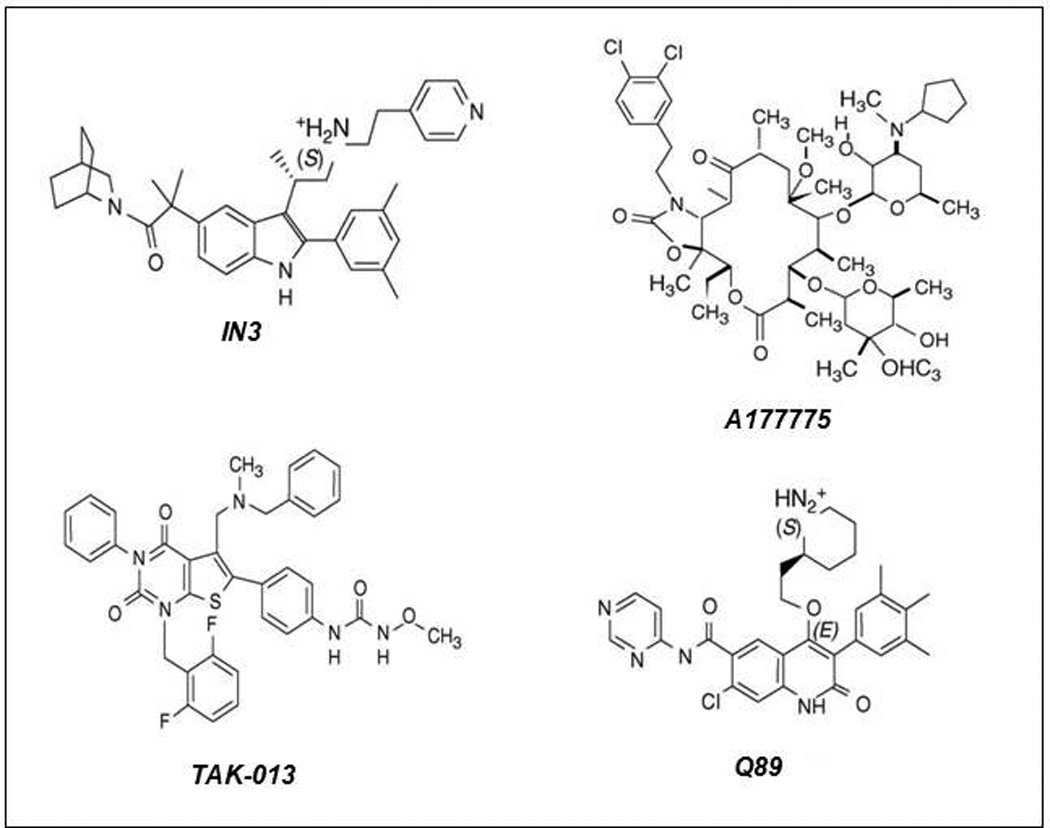
Structural alterations provoked by mutations or genetic variations in the gene sequence of G protein-coupled receptors (GPCRs) may lead to abnormal function of the receptor molecule. Frequently, this leads to disease. While some mutations lead to changes in domains involved in agonist binding, receptor activation, or coupling to effectors, others may cause misfolding and lead to retention/degradation of the protein molecule by the quality control system of the cell. Several strategies, including genetic, chemical, and pharmacological approaches, have been shown to rescue function of trafficking-defective misfolded GPCRs. Among these, pharmacological strategies offer the most promising therapeutic tool to promote proper trafficking of misfolded proteins to the plasma membrane (PM). Pharmacological chaperones or "pharmacoperones" are small compounds that permeate the PM, enter cells, and bind selectively to misfolded proteins and correct folding allowing routing of the target protein to the PM, where the receptor may bind and respond to agonist stimulation. In this review, we describe new therapeutic opportunities based on mislocalization of otherwise functional human gonadotropin-releasing hormone receptors. This particular receptor is highly sensitive to single changes in chemical charge, and its intracellular traffic is delicately balanced between expression at the PM or retention/degradation in the endoplasmic reticulum; it is, therefore, a particularly instructive model to understand both the protein routing and the molecular mechanisms, whereby pharmacoperones rescue misfolded intermediates or conformationally defective receptors.
Copyright © 2011 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures
References
-
- Aittomaki K, Lucena JL, Pakarinen P, Sistonen P, Tapanainen J, Gromoll J, et al. Mutation in the follicle-stimulating hormone receptor gene causes hereditary hypergonadotropic ovarian failure. Cell. 1995;82(6):959–968. - PubMed
-
- Amaral MD. Therapy through chaperones: sense or antisense? Cystic fibrosis as a model disease. J Inherit Metab Dis. 2006;29(2–3):477–487. - PubMed
-
- Amiel J, Sproat-Emison E, Garcia-Barcelo M, Lantieri F, Burzynski G, Borrego S, et al. Hirschsprung disease, associated syndromes and genetics: a review. J Med Genet. 2008;45(1):1–14. - PubMed
-
- Andersson H, D'Antona AM, Kendall DA, Von Heijne G, Chin CN. Membrane assembly of the cannabinoid receptor 1: impact of a long N-terminal tail. Mol Pharmacol. 2003;64(3):570–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
