

Loss of ribosomal RNA modification causes developmental defects in zebrafish
- PMID: 21908402
- PMCID: PMC3245925
- DOI: 10.1093/nar/gkr700
Loss of ribosomal RNA modification causes developmental defects in zebrafish
Abstract
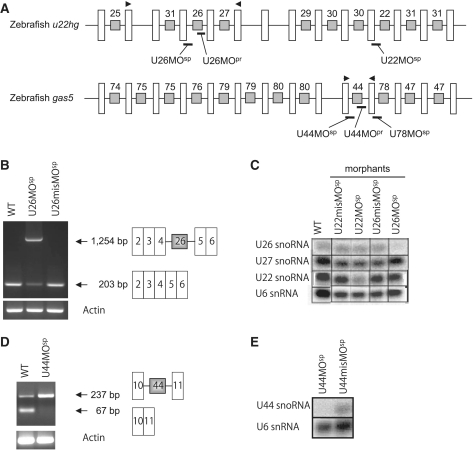
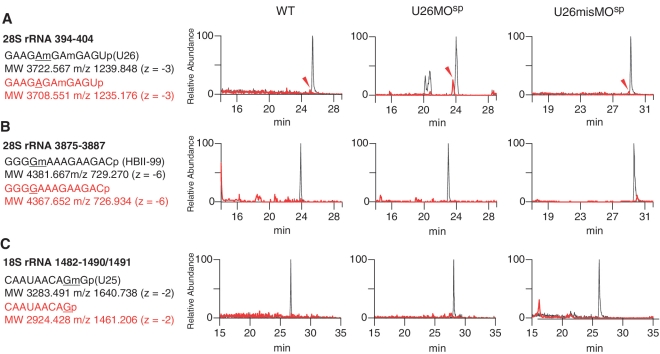
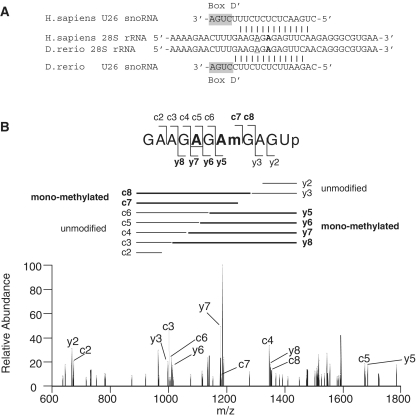
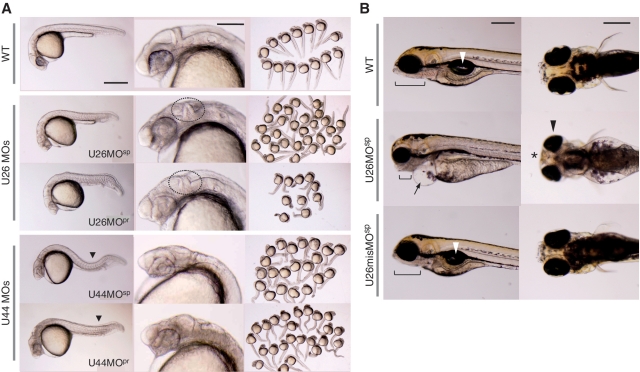
Non-coding RNAs (ncRNAs) play key roles in diverse cellular activities, and efficient ncRNA function requires extensive posttranscriptional nucleotide modifications. Small nucleolar RNAs (snoRNAs) are a group of ncRNAs that guide the modification of specific nucleotides in ribosomal RNAs (rRNAs) and small nuclear RNAs. To investigate the physiological relevance of rRNA modification in vertebrates, we suppressed the expression of three snoRNAs (U26, U44 and U78), either by disrupting the host gene splicing or by inhibiting the snoRNA precursor processing, and analyzed the consequences of snoRNA loss-of-function in zebrafish. Using a highly sensitive mass spectrometric analysis, we found that decreased snoRNA expression reduces the snoRNA-guided methylation of the target nucleotides. Impaired rRNA modification, even at a single site, led to severe morphological defects and embryonic lethality in zebrafish, which suggests that rRNA modifications play an essential role in vertebrate development. This study highlights the importance of posttranscriptional modifications and their role in ncRNA function in higher eukaryotes.
Figures
References
-
- Suzuki T. Biosynthesis and function of tRNA wobble modifications. In: Grosjean H, editor. Fine-Tuning of RNA Functions by Modification and Editing. Berlin Heidelberg: Springer-Verlag; 2005. pp. 23–69.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
