

Host-derived adiponectin is tumor-suppressive and a novel therapeutic target for multiple myeloma and the associated bone disease
- PMID: 21908434
- PMCID: PMC3228502
- DOI: 10.1182/blood-2011-01-330407
Host-derived adiponectin is tumor-suppressive and a novel therapeutic target for multiple myeloma and the associated bone disease
Abstract
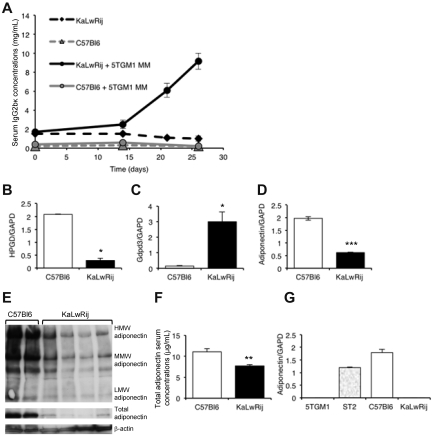
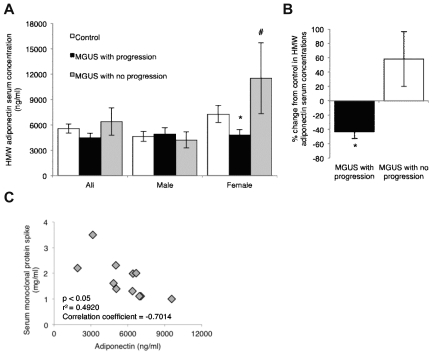
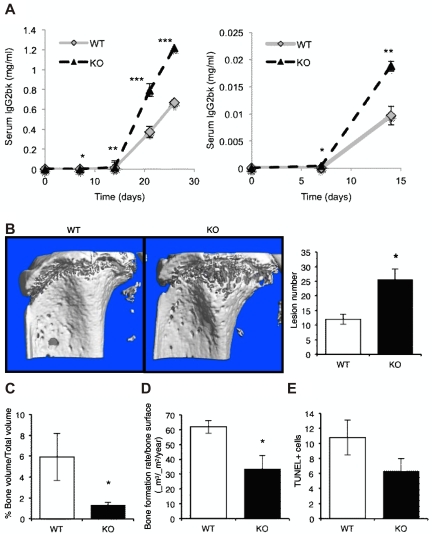
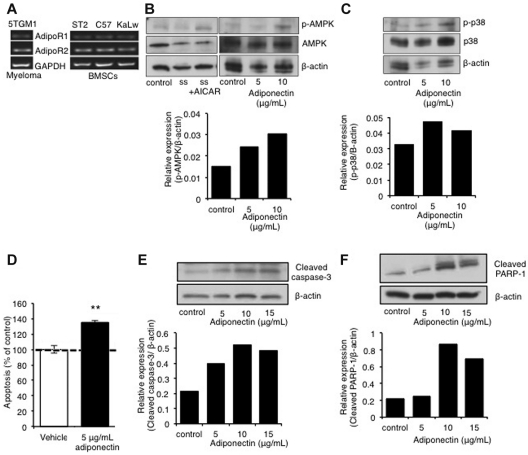
The contributions of the host microenvironment to the pathogenesis of multiple myeloma, including progression from the non-malignant disorder monoclonal gammopathy of undetermined significance, are poorly understood. In the present study, microarray analysis of a murine model requiring a unique host microenvironment for myeloma development identified decreased host-derived adiponectin compared with normal mice. In support, clinical analysis revealed decreased serum adiponectin concentrations in monoclonal gammopathy of undetermined significance patients who subsequently progressed to myeloma. We investigated the role of adiponectin in myeloma pathogenesis and as a treatment approach, using both mice deficient in adiponectin and pharmacologic enhancement of circulating adiponectin. Increased tumor burden and bone disease were observed in myeloma-bearing adiponectin-deficient mice, and adiponectin was found to induce myeloma cell apoptosis. The apolipoprotein peptide mimetic L-4F was used for pharmacologic enhancement of adiponectin. L-4F reduced tumor burden, increased survival of myeloma-bearing mice, and prevented myeloma bone disease. Collectively, our studies have identified a novel mechanism whereby decreased host-derived adiponectin promotes myeloma tumor growth and osteolysis. Furthermore, we have established the potential therapeutic benefit of increasing adiponectin for the treatment of myeloma and the associated bone disease.
Figures
Comment in
-
Adipokines in MM: time to trim the fat.Blood. 2011 Nov 24;118(22):5716-7. doi: 10.1182/blood-2011-10-379834. Blood. 2011. PMID: 22123904 No abstract available.
References
-
- Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60(5):277–300. - PubMed
-
- Radl J, de Glopper E, Schuit HER, Zurcher C. Idiopathic paraprotienemia: II. Transplantation of the paraprotein-producing clone from old to young C587Bl/KaLwRij mice. J Immunol. 1979;122(2):609–613. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
