

Preference as a function of active interresponse times: a test of the active time model
- PMID: 21909165
- PMCID: PMC3168888
- DOI: 10.1901/jeab.2011.96-215
Preference as a function of active interresponse times: a test of the active time model
Abstract
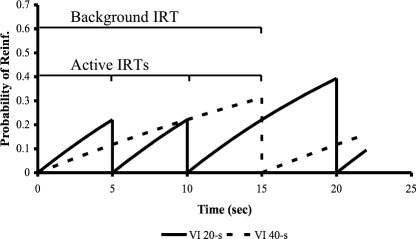
In this article, we describe a test of the active time model for concurrent variable interval (VI) choice. The active time model (ATM) suggests that the time since the most recent response is one of the variables controlling choice in concurrent VI VI schedules of reinforcement. In our experiment, pigeons were trained in a multiple concurrent similar to that employed by Belke (1992), with VI 20-s and VI 40-s schedules in one component, and VI 40-s and VI 80-s schedules in the other component. However, rather than use a free-operant design, we used a discrete-trial procedure that restricted interresponse times to a range of 0.5-9.0 s. After 45 sessions of training, unreinforced probe periods were mixed with reinforced training periods. These probes paired the two stimuli associated with the VI 40-s schedules. Further, the probes were defined such that during their occurrence, interresponse times were either "short" (0.5-3.0 s) or "long" (7.5-9.0 s). All pigeons showed a preference for the stimulus associated with the relatively rich VI 40-s schedule--a result mirroring that of Belke. We also observed, though, that this preference was more extreme during long probes than during short probes--a result predicted by ATM.
Keywords: Belke (1992); active time model; interresponse times; pigeons; stay/switch model; variable-intervals.
Figures





Similar articles
-
The active time model of concurrent choice.PLoS One. 2024 May 21;19(5):e0301173. doi: 10.1371/journal.pone.0301173. eCollection 2024. PLoS One. 2024. PMID: 38771859 Free PMC article.
-
An application of the active time model to multiple concurrent variable-interval schedules.Behav Processes. 2009 Jun;81(2):250-5. doi: 10.1016/j.beproc.2008.10.014. Epub 2008 Nov 6. Behav Processes. 2009. PMID: 19038316
-
A further application of the active time model to multiple concurrent variable-interval schedules.Behav Processes. 2010 May;84(1):470-5. doi: 10.1016/j.beproc.2009.09.006. Epub 2009 Oct 6. Behav Processes. 2010. PMID: 19815057
-
Arousal, changeover responses, and preference in concurrent schedules.J Exp Anal Behav. 2003 Nov;80(3):261-72. doi: 10.1901/jeab.2003.80-261. J Exp Anal Behav. 2003. PMID: 14964707 Free PMC article.
-
The copyist model of response emission.Psychon Bull Rev. 2012 Oct;19(5):759-78. doi: 10.3758/s13423-012-0267-1. Psychon Bull Rev. 2012. PMID: 22673925 Review.
Cited by
-
Regularities in responding during performance of a complex choice task.Learn Behav. 2015 Dec;43(4):323-41. doi: 10.3758/s13420-015-0182-1. Learn Behav. 2015. PMID: 26077440
-
The active time model of concurrent choice.PLoS One. 2024 May 21;19(5):e0301173. doi: 10.1371/journal.pone.0301173. eCollection 2024. PLoS One. 2024. PMID: 38771859 Free PMC article.
-
Resistance to extinction and relapse in combined stimulus contexts.J Exp Anal Behav. 2012 Sep;98(2):169-89. doi: 10.1901/jeab.2012.98-169. J Exp Anal Behav. 2012. PMID: 23008521 Free PMC article.
References
-
- Belke T.W. Stimulus preference and the transitivity of preference. Animal Learning & Behavior. 1992;20:401–406.
-
- Brown E, Cleaveland J.M. An application of the active time model to multiple concurrent variable-interval schedules. Behavioural Processes. 2009;81:250–255. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
