

Influenza virus ribonucleoprotein complexes gain preferential access to cellular export machinery through chromatin targeting
- PMID: 21909257
- PMCID: PMC3164630
- DOI: 10.1371/journal.ppat.1002187
Influenza virus ribonucleoprotein complexes gain preferential access to cellular export machinery through chromatin targeting
Abstract
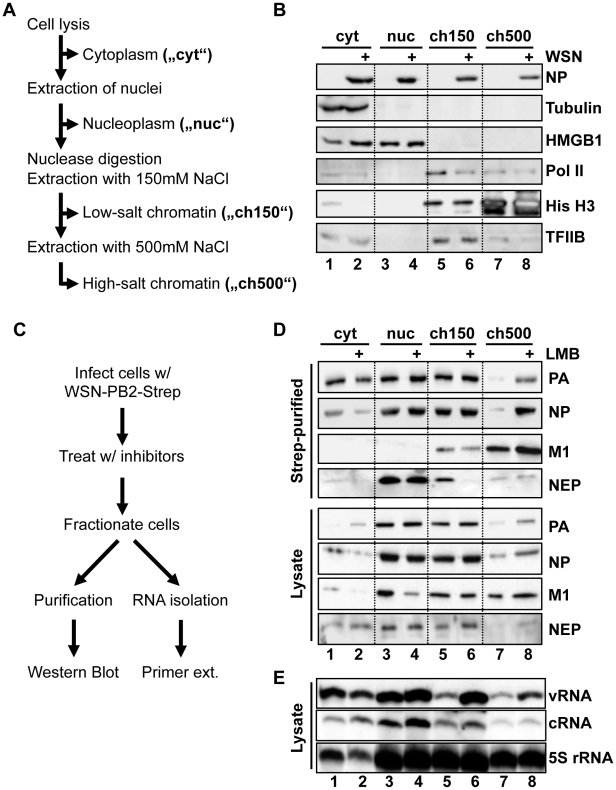
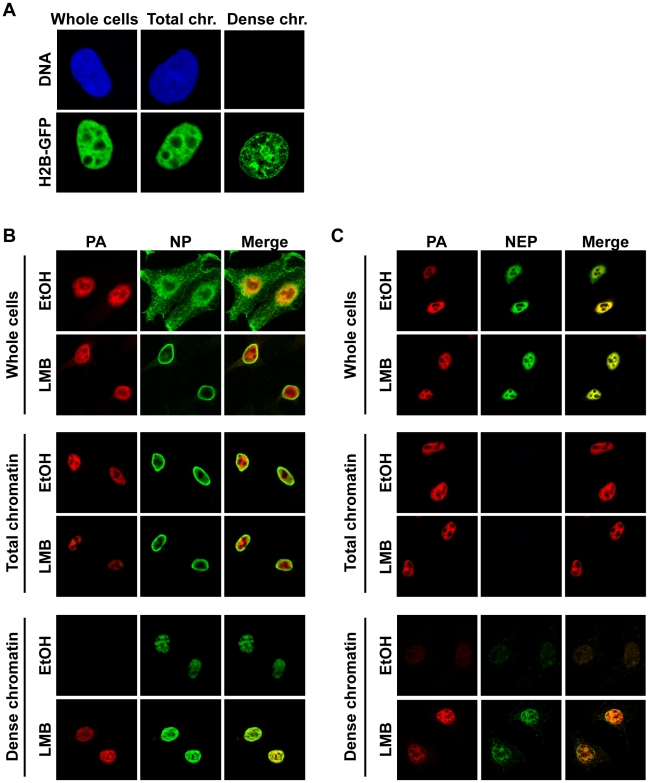
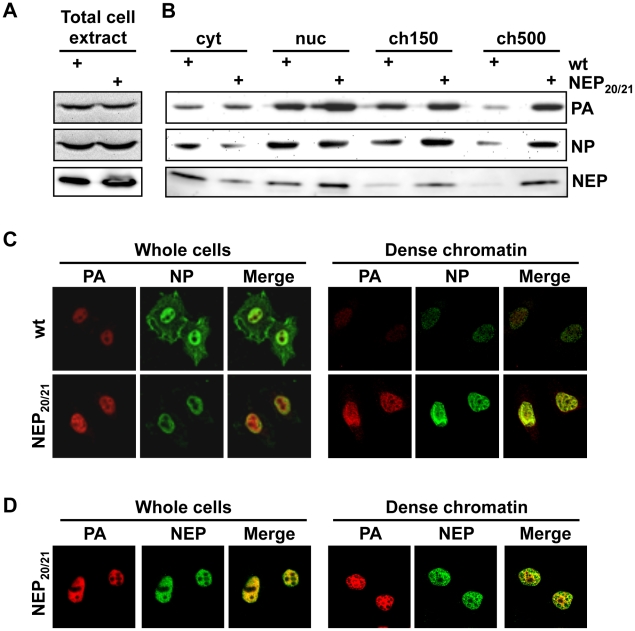
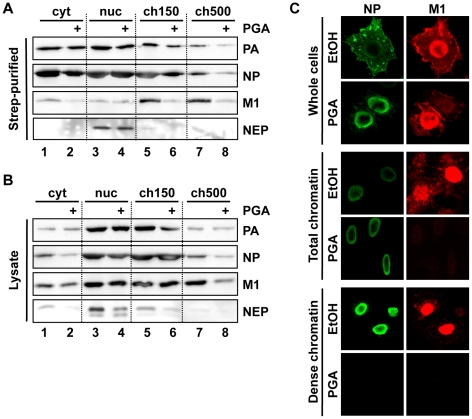
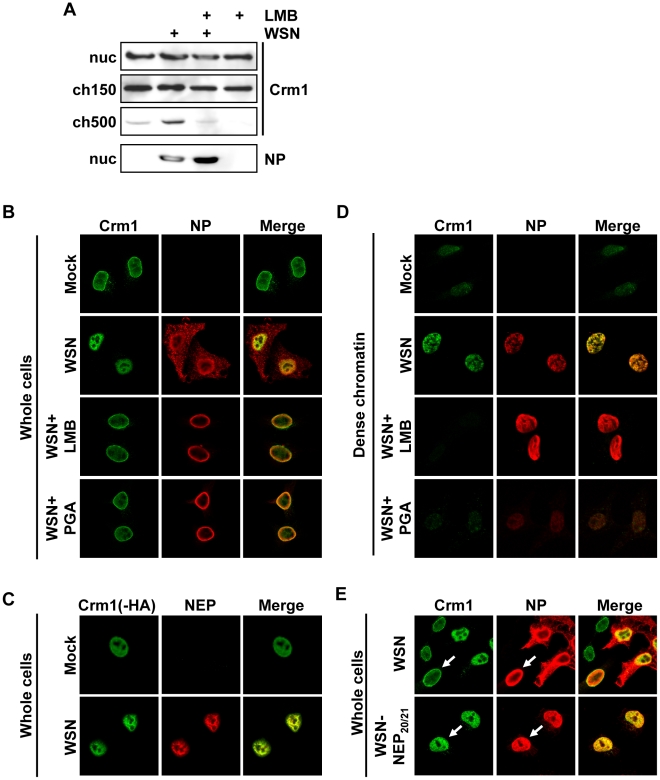
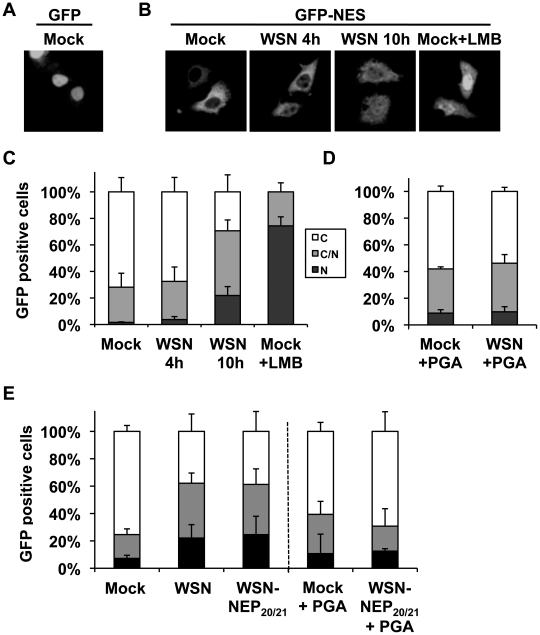
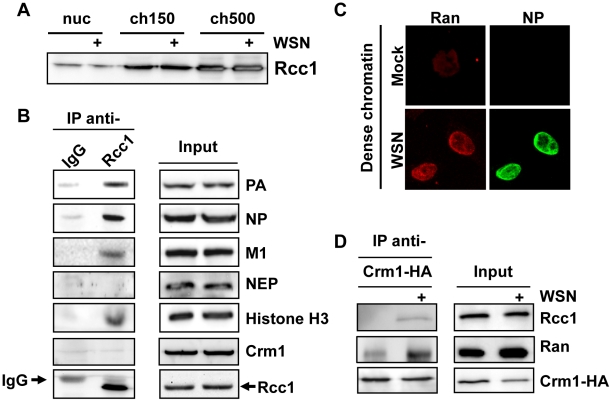
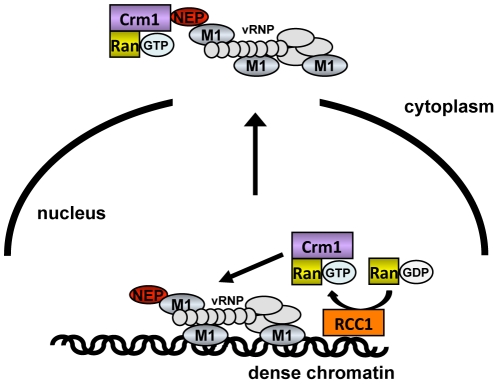
In contrast to most RNA viruses, influenza viruses replicate their genome in the nucleus of infected cells. As a result, newly-synthesized vRNA genomes, in the form of viral ribonucleoprotein complexes (vRNPs), must be exported to the cytoplasm for productive infection. To characterize the composition of vRNP export complexes and their interplay with the nucleus of infected cells, we affinity-purified tagged vRNPs from biochemically fractionated infected nuclei. After treatment of infected cells with leptomycin B, a potent inhibitor of Crm1-mediated export, we isolated vRNP export complexes which, unexpectedly, were tethered to the host-cell chromatin with very high affinity. At late time points of infection, the cellular export receptor Crm1 also accumulated at the same regions of the chromatin as vRNPs, which led to a decrease in the export of other nuclear Crm1 substrates from the nucleus. Interestingly, chromatin targeting of vRNP export complexes brought them into association with Rcc1, the Ran guanine exchange factor responsible for generating RanGTP and driving Crm1-dependent nuclear export. Thus, influenza viruses gain preferential access to newly-generated host cell export machinery by targeting vRNP export complexes at the sites of Ran regeneration.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Selective incorporation of vRNP into influenza A virions determined by its specific interaction with M1 protein.Virology. 2017 May;505:23-32. doi: 10.1016/j.virol.2017.02.008. Epub 2017 Feb 17. Virology. 2017. PMID: 28219018 Free PMC article.
-
Inhibition of RAN attenuates influenza a virus replication and nucleoprotein nuclear export.Emerg Microbes Infect. 2024 Dec;13(1):2387910. doi: 10.1080/22221751.2024.2387910. Epub 2024 Aug 12. Emerg Microbes Infect. 2024. PMID: 39087696 Free PMC article.
-
Inhibition of nuclear export of ribonucleoprotein complexes of influenza virus by leptomycin B.Virus Res. 2001 Sep;77(1):31-42. doi: 10.1016/s0168-1702(01)00263-5. Virus Res. 2001. PMID: 11451485
-
Host Cell Factors That Interact with Influenza Virus Ribonucleoproteins.Cold Spring Harb Perspect Med. 2021 Nov 1;11(11):a038307. doi: 10.1101/cshperspect.a038307. Cold Spring Harb Perspect Med. 2021. PMID: 32988980 Free PMC article. Review.
-
Biogenesis, assembly, and export of viral messenger ribonucleoproteins in the influenza A virus infected cell.RNA Biol. 2013 Aug;10(8):1274-82. doi: 10.4161/rna.25356. Epub 2013 Jun 17. RNA Biol. 2013. PMID: 23807439 Free PMC article. Review.
Cited by
-
Comparative analysis of retroviral Gag-host cell interactions: focus on the nuclear interactome.bioRxiv [Preprint]. 2024 Mar 6:2024.01.18.575255. doi: 10.1101/2024.01.18.575255. bioRxiv. 2024. Update in: Retrovirology. 2024 Jun 19;21(1):13. doi: 10.1186/s12977-024-00645-y. PMID: 38293010 Free PMC article. Updated. Preprint.
-
Selective incorporation of vRNP into influenza A virions determined by its specific interaction with M1 protein.Virology. 2017 May;505:23-32. doi: 10.1016/j.virol.2017.02.008. Epub 2017 Feb 17. Virology. 2017. PMID: 28219018 Free PMC article.
-
Inhibition of RAN attenuates influenza a virus replication and nucleoprotein nuclear export.Emerg Microbes Infect. 2024 Dec;13(1):2387910. doi: 10.1080/22221751.2024.2387910. Epub 2024 Aug 12. Emerg Microbes Infect. 2024. PMID: 39087696 Free PMC article.
-
Infectious Salmon Anemia Virus Infectivity Is Determined by Multiple Segments with an Important Contribution from Segment 5.Viruses. 2022 Mar 18;14(3):631. doi: 10.3390/v14030631. Viruses. 2022. PMID: 35337038 Free PMC article.
-
A comprehensive map of the influenza A virus replication cycle.BMC Syst Biol. 2013 Oct 2;7:97. doi: 10.1186/1752-0509-7-97. BMC Syst Biol. 2013. PMID: 24088197 Free PMC article.
References
-
- Martin K, Helenius A. Nuclear transport of influenza virus ribonucleoproteins: the viral matrix protein (M1) promotes export and inhibits import. Cell. 1991;67:117–130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
