

Plasmodium protease ROM1 is important for proper formation of the parasitophorous vacuole
- PMID: 21909259
- PMCID: PMC3164628
- DOI: 10.1371/journal.ppat.1002197
Plasmodium protease ROM1 is important for proper formation of the parasitophorous vacuole
Abstract
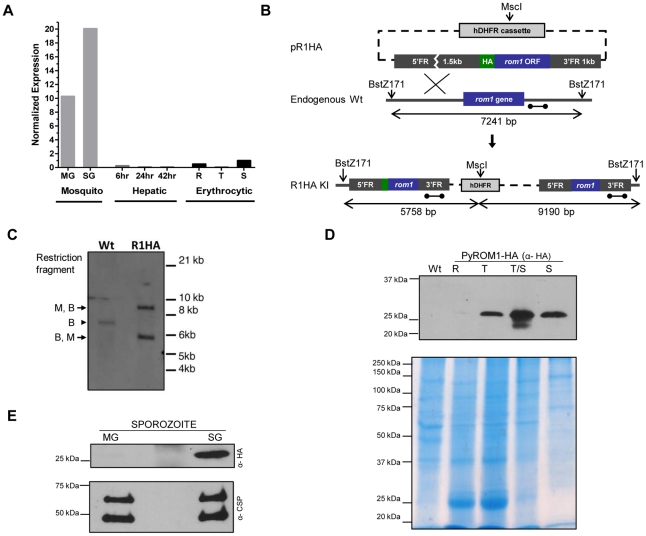
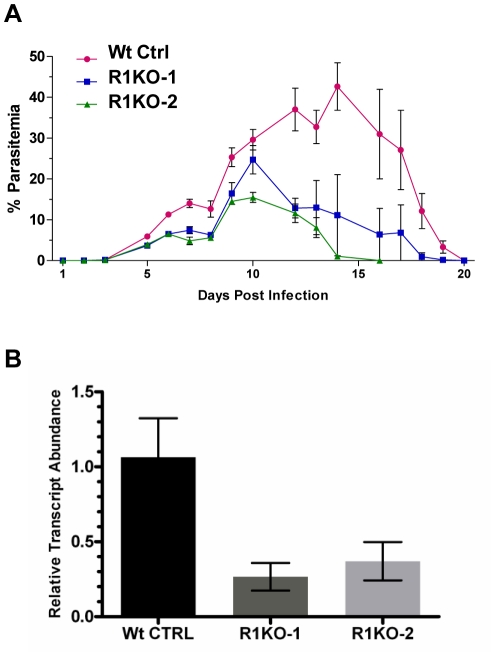
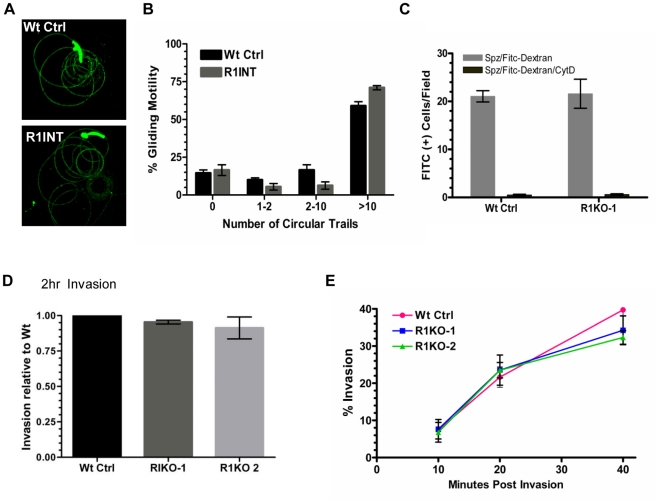
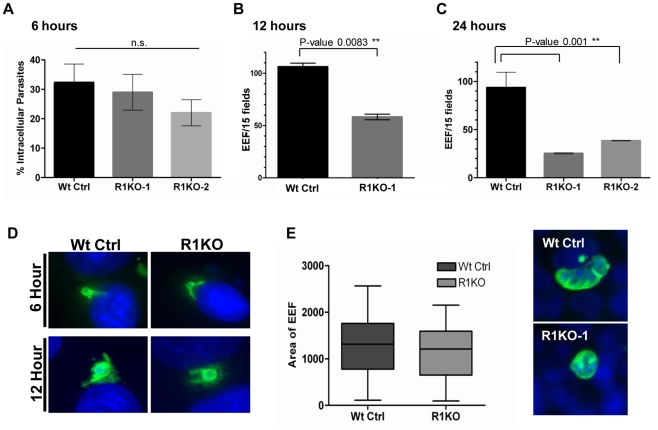
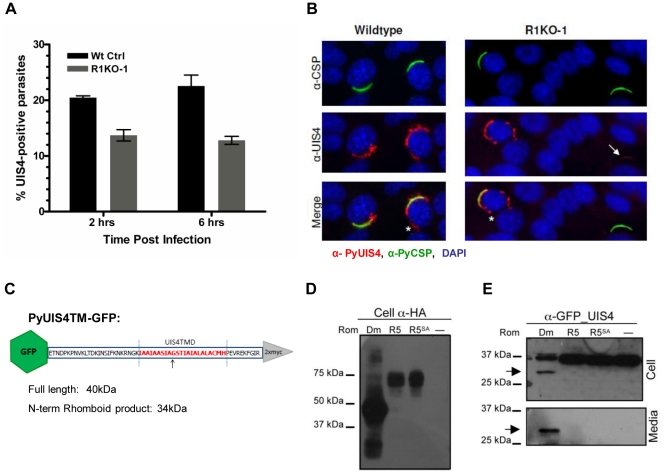
Apicomplexans are obligate intracellular parasites that invade host cells by an active process leading to the formation of a non-fusogenic parasitophorous vacuole (PV) where the parasite replicates within the host cell. The rhomboid family of proteases cleaves substrates within their transmembrane domains and has been implicated in the invasion process. Although its exact function is unknown, Plasmodium ROM1 is hypothesized to play a role during invasion based on its microneme localization and its ability to cleave essential invasion adhesins. Using the rodent malaria model, Plasmodium yoelii, we carried out detailed quantitative analysis of pyrom1 deficient parasites during the Plasmodium lifecycle. Pyrom1(-) parasites are attenuated during erythrocytic and hepatic stages but progress normally through the mosquito vector with normal counts of oocyst and salivary gland sporozoites. Pyrom1 steady state mRNA levels are upregulated 20-fold in salivary gland sporozoites compared to blood stages. We show that pyrom1(-) sporozoites are capable of gliding motility and traversing host cells normally. Wildtype and pyrom1(-) sporozoites do not differ in the rate of entry into Hepa1-6 hepatocytes. Within the first twelve hours of hepatic development, however, only 50% pyrom1(-) parasites have developed into exoerythrocytic forms. Immunofluorescence microscopy using the PVM marker UIS4 and transmission electron microscopy reveal that the PV of a significant fraction of pyrom1(-) parasites are morphologically aberrant shortly after invasion. We propose a novel function for PyROM1 as a protease that promotes proper PV modification to allow parasite development and replication in a suitable environment within the mammalian host.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Plasmodium yoelii sporozoites with simultaneous deletion of P52 and P36 are completely attenuated and confer sterile immunity against infection.Infect Immun. 2007 Aug;75(8):3758-68. doi: 10.1128/IAI.00225-07. Epub 2007 May 21. Infect Immun. 2007. PMID: 17517871 Free PMC article.
-
Malaria Sporozoites Traverse Host Cells within Transient Vacuoles.Cell Host Microbe. 2015 Nov 11;18(5):593-603. doi: 10.1016/j.chom.2015.10.006. Cell Host Microbe. 2015. PMID: 26607162
-
The Micronemal Plasmodium Proteins P36 and P52 Act in Concert to Establish the Replication-Permissive Compartment Within Infected Hepatocytes.Front Cell Infect Microbiol. 2018 Nov 27;8:413. doi: 10.3389/fcimb.2018.00413. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 30547015 Free PMC article.
-
Plasmodium sporozoites on the move: Switching from cell traversal to productive invasion of hepatocytes.Mol Microbiol. 2021 May;115(5):870-881. doi: 10.1111/mmi.14645. Epub 2020 Dec 5. Mol Microbiol. 2021. PMID: 33191548 Free PMC article. Review.
-
Molecular make-up of the Plasmodium parasitophorous vacuolar membrane.Int J Med Microbiol. 2012 Oct;302(4-5):179-86. doi: 10.1016/j.ijmm.2012.07.011. Epub 2012 Aug 13. Int J Med Microbiol. 2012. PMID: 22898489 Review.
Cited by
-
Malaria parasite pre-erythrocytic infection: preparation meets opportunity.Cell Microbiol. 2012 Mar;14(3):316-24. doi: 10.1111/j.1462-5822.2011.01734.x. Epub 2012 Jan 9. Cell Microbiol. 2012. PMID: 22151703 Free PMC article. Review.
-
Entamoeba histolytica rhomboid protease 1 has a role in migration and motility as validated by two independent genetic approaches.Exp Parasitol. 2015 Jul;154:33-42. doi: 10.1016/j.exppara.2015.04.004. Epub 2015 Apr 15. Exp Parasitol. 2015. PMID: 25889553 Free PMC article.
-
Torins are potent antimalarials that block replenishment of Plasmodium liver stage parasitophorous vacuole membrane proteins.Proc Natl Acad Sci U S A. 2013 Jul 23;110(30):E2838-47. doi: 10.1073/pnas.1306097110. Epub 2013 Jul 8. Proc Natl Acad Sci U S A. 2013. PMID: 23836641 Free PMC article.
-
Plasmodium berghei Δp52&p36 parasites develop independent of a parasitophorous vacuole membrane in Huh-7 liver cells.PLoS One. 2012;7(12):e50772. doi: 10.1371/journal.pone.0050772. Epub 2012 Dec 5. PLoS One. 2012. PMID: 23227206 Free PMC article.
-
Post-Translational Modifications of Proteins of Malaria Parasites during the Life Cycle.Int J Mol Sci. 2024 Jun 2;25(11):6145. doi: 10.3390/ijms25116145. Int J Mol Sci. 2024. PMID: 38892332 Free PMC article. Review.
References
-
- Baum J, Gilberger TW, Frischknecht F, Meissner M. Host-cell invasion by malaria parasites: insights from Plasmodium and Toxoplasma. Trends Parasitol. 2008;24:557–563. - PubMed
-
- Huynh MH, Harper JM, Carruthers VB. Preparing for an invasion: charting the pathway of adhesion proteins to Toxoplasma micronemes. Parasitol Res. 2006;98:389–395. - PubMed
-
- Lebrun M, Michelin A, El Hajj H, Poncet J, Bradley PJ, et al. The rhoptry neck protein RON4 re-localizes at the moving junction during Toxoplasma gondii invasion. Cell Microbiol. 2005;7:1823–1833. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
