

The motility of a human parasite, Toxoplasma gondii, is regulated by a novel lysine methyltransferase
- PMID: 21909263
- PMCID: PMC3164638
- DOI: 10.1371/journal.ppat.1002201
The motility of a human parasite, Toxoplasma gondii, is regulated by a novel lysine methyltransferase
Abstract
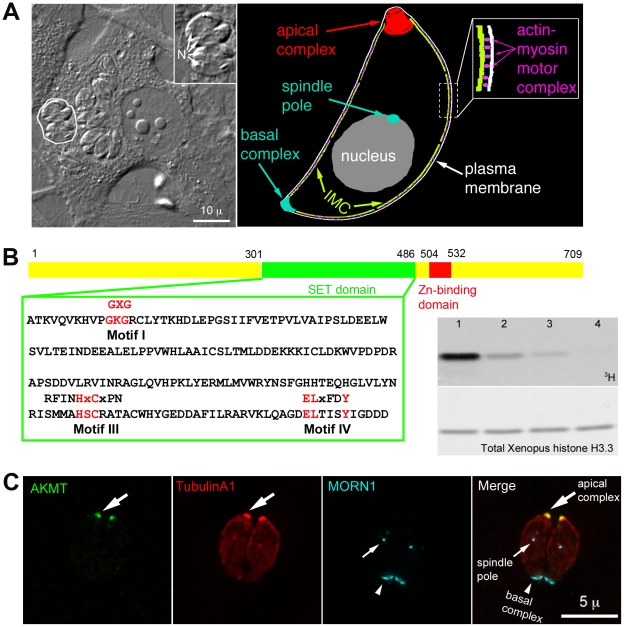
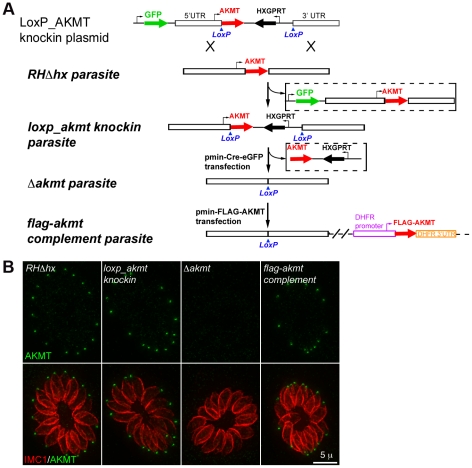
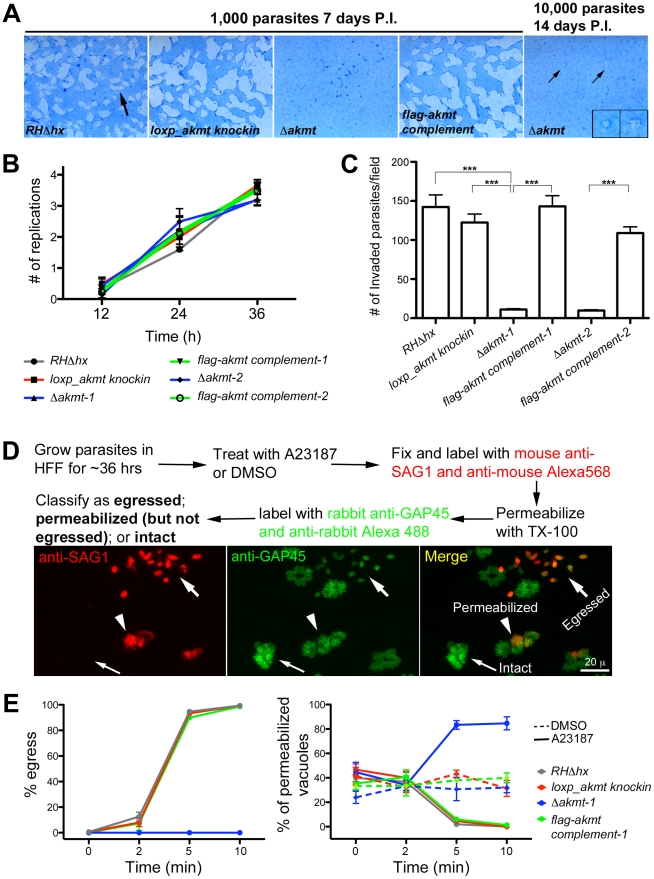
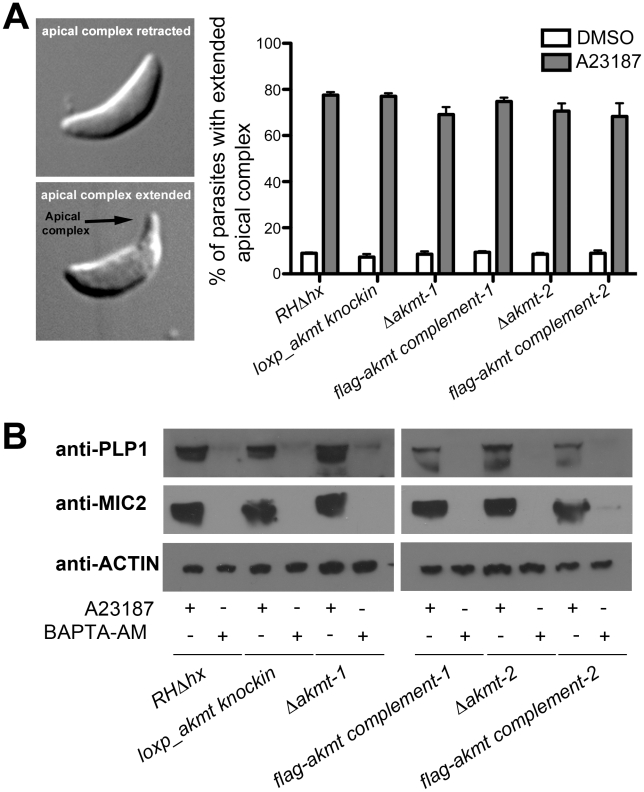
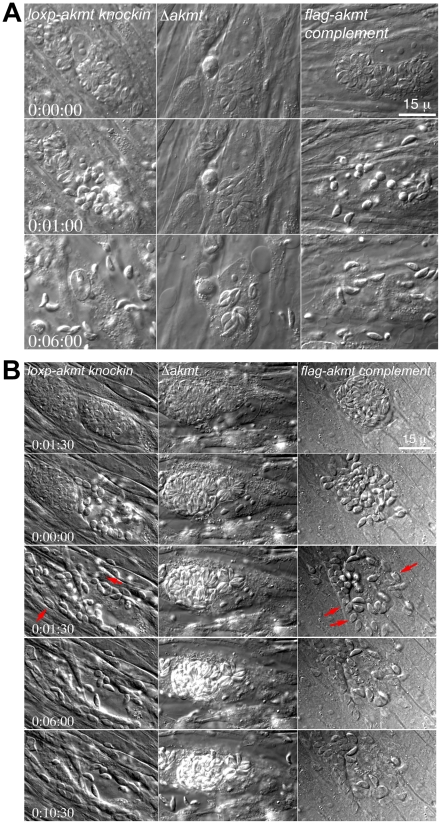
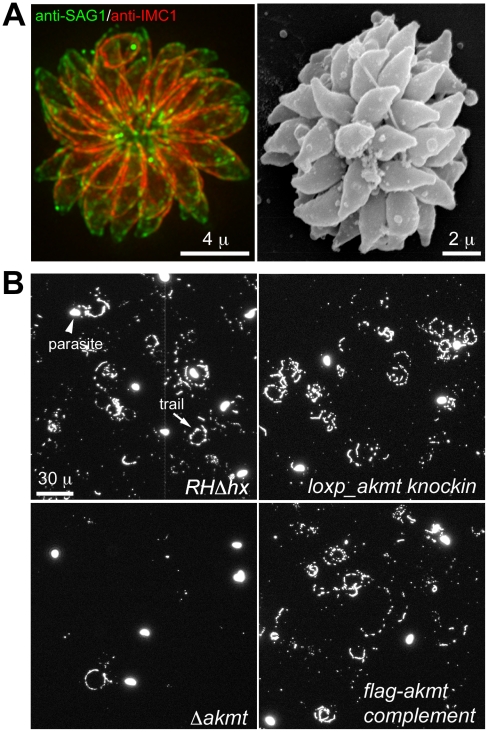
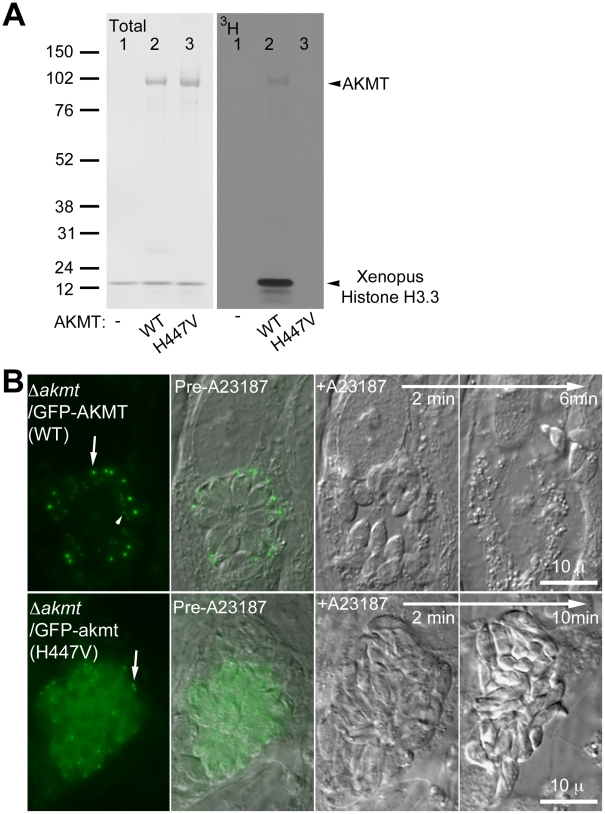
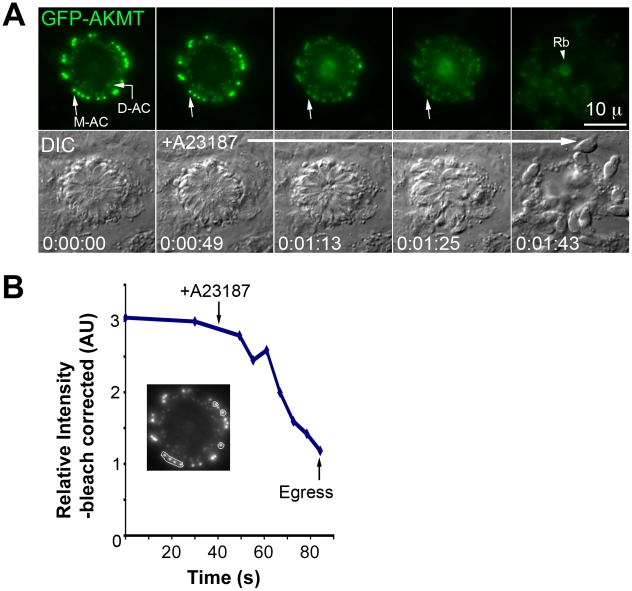
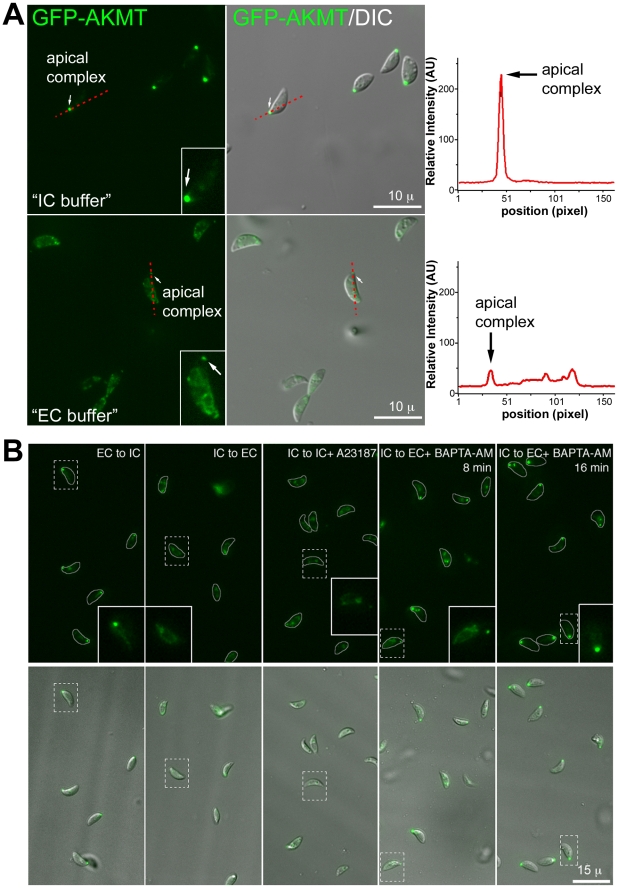
Protozoa in the phylum Apicomplexa are a large group of obligate intracellular parasites. Toxoplasma gondii and other apicomplexan parasites, such as Plasmodium falciparum, cause diseases by reiterating their lytic cycle, comprising host cell invasion, parasite replication, and parasite egress. The successful completion of the lytic cycle requires that the parasite senses changes in its environment and switches between the non-motile (for intracellular replication) and motile (for invasion and egress) states appropriately. Although the signaling pathway that regulates the motile state switch is critical to the pathogenesis of the diseases caused by these parasites, it is not well understood. Here we report a previously unknown mechanism of regulating the motility activation in Toxoplasma, mediated by a protein lysine methyltransferase, AKMT (for Apical complex lysine (K) methyltransferase). AKMT depletion greatly inhibits activation of motility, compromises parasite invasion and egress, and thus severely impairs the lytic cycle. Interestingly, AKMT redistributes from the apical complex to the parasite body rapidly in the presence of egress-stimulating signals that increase [Ca²⁺] in the parasite cytoplasm, suggesting that AKMT regulation of parasite motility might be accomplished by the precise temporal control of its localization in response to environmental changes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Direct measurement of cortical force generation and polarization in a living parasite.Mol Biol Cell. 2017 Jul 7;28(14):1912-1923. doi: 10.1091/mbc.E16-07-0518. Epub 2017 Feb 16. Mol Biol Cell. 2017. PMID: 28209732 Free PMC article.
-
Identification of functional modules of AKMT, a novel lysine methyltransferase regulating the motility of Toxoplasma gondii.Mol Biochem Parasitol. 2013 May;189(1-2):43-53. doi: 10.1016/j.molbiopara.2013.05.004. Epub 2013 May 17. Mol Biochem Parasitol. 2013. PMID: 23685344 Free PMC article.
-
Structure of a Novel Dimeric SET Domain Methyltransferase that Regulates Cell Motility.J Mol Biol. 2018 Oct 19;430(21):4209-4229. doi: 10.1016/j.jmb.2018.08.017. Epub 2018 Aug 24. J Mol Biol. 2018. PMID: 30148980 Free PMC article.
-
Calcium signaling and the lytic cycle of the Apicomplexan parasite Toxoplasma gondii.Biochim Biophys Acta Mol Cell Res. 2018 Nov;1865(11 Pt B):1846-1856. doi: 10.1016/j.bbamcr.2018.08.004. Epub 2018 Aug 10. Biochim Biophys Acta Mol Cell Res. 2018. PMID: 30992126 Free PMC article. Review.
-
Signaling Cascades Governing Entry into and Exit from Host Cells by Toxoplasma gondii.Annu Rev Microbiol. 2019 Sep 8;73:579-599. doi: 10.1146/annurev-micro-020518-120235. Annu Rev Microbiol. 2019. PMID: 31500539 Review.
Cited by
-
The apical annuli of Toxoplasma gondii are composed of coiled-coil and signalling proteins embedded in the inner membrane complex sutures.Cell Microbiol. 2020 Jan;22(1):e13112. doi: 10.1111/cmi.13112. Epub 2019 Sep 10. Cell Microbiol. 2020. PMID: 31470470 Free PMC article.
-
Loss of a doublecortin (DCX)-domain protein causes structural defects in a tubulin-based organelle of Toxoplasma gondii and impairs host-cell invasion.Mol Biol Cell. 2017 Feb 1;28(3):411-428. doi: 10.1091/mbc.E16-08-0587. Epub 2016 Dec 8. Mol Biol Cell. 2017. PMID: 27932494 Free PMC article.
-
Direct measurement of cortical force generation and polarization in a living parasite.Mol Biol Cell. 2017 Jul 7;28(14):1912-1923. doi: 10.1091/mbc.E16-07-0518. Epub 2017 Feb 16. Mol Biol Cell. 2017. PMID: 28209732 Free PMC article.
-
Temporal and thermal profiling of the Toxoplasma proteome implicates parasite Protein Phosphatase 1 in the regulation of Ca2+-responsive pathways.Elife. 2022 Aug 17;11:e80336. doi: 10.7554/eLife.80336. Elife. 2022. PMID: 35976251 Free PMC article.
-
Essential function of the alveolin network in the subpellicular microtubules and conoid assembly in Toxoplasma gondii.Elife. 2020 May 7;9:e56635. doi: 10.7554/eLife.56635. Elife. 2020. PMID: 32379047 Free PMC article.
References
-
- Levine ND. Progress in taxonomy of the Apicomplexan protozoa. J Protozool. 1988;35:518–520. - PubMed
-
- Dobrowolski JM, Carruthers VB, Sibley LD. Participation of myosin in gliding motility and host cell invasion by Toxoplasma gondii. Mol Microbiol. 1997;26:163–173. - PubMed
-
- Lingelbach K, Joiner KA. The parasitophorous vacuole membrane surrounding Plasmodium and Toxoplasma: an unusual compartment in infected cells. J Cell Sci. 1998;111:1467–1475. - PubMed
-
- Sheffield HG, Melton ML. The fine structure and reproduction of Toxoplasma gondii. J Parasitol. 1968;54:209–226. - PubMed
-
- Sinai AP, Joiner KA. Safe haven: the cell biology of nonfusogenic pathogen vacuoles. Annu Rev Microbiol. 1997;51:415–462. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
