

Adult subependymal neural precursors, but not differentiated cells, undergo rapid cathodal migration in the presence of direct current electric fields
- PMID: 21909360
- PMCID: PMC3166127
- DOI: 10.1371/journal.pone.0023808
Adult subependymal neural precursors, but not differentiated cells, undergo rapid cathodal migration in the presence of direct current electric fields
Abstract
Background: The existence of neural stem and progenitor cells (together termed neural precursor cells) in the adult mammalian brain has sparked great interest in utilizing these cells for regenerative medicine strategies. Endogenous neural precursors within the adult forebrain subependyma can be activated following injury, resulting in their proliferation and migration toward lesion sites where they differentiate into neural cells. The administration of growth factors and immunomodulatory agents following injury augments this activation and has been shown to result in behavioural functional recovery following stroke.
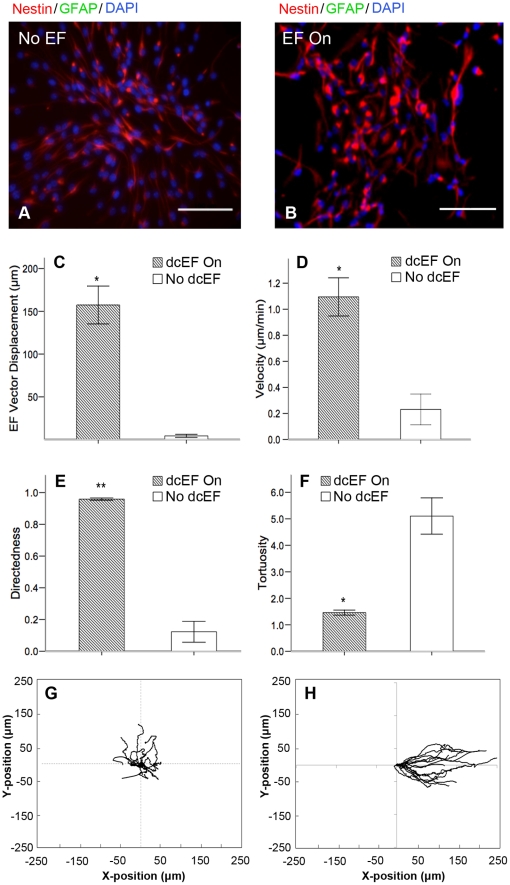
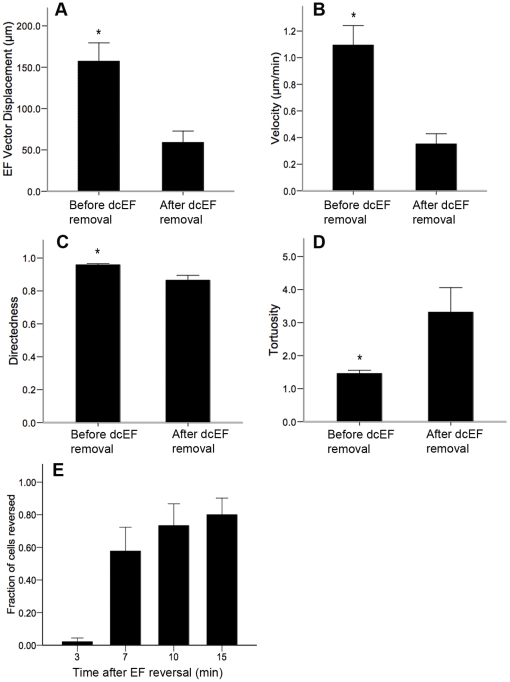
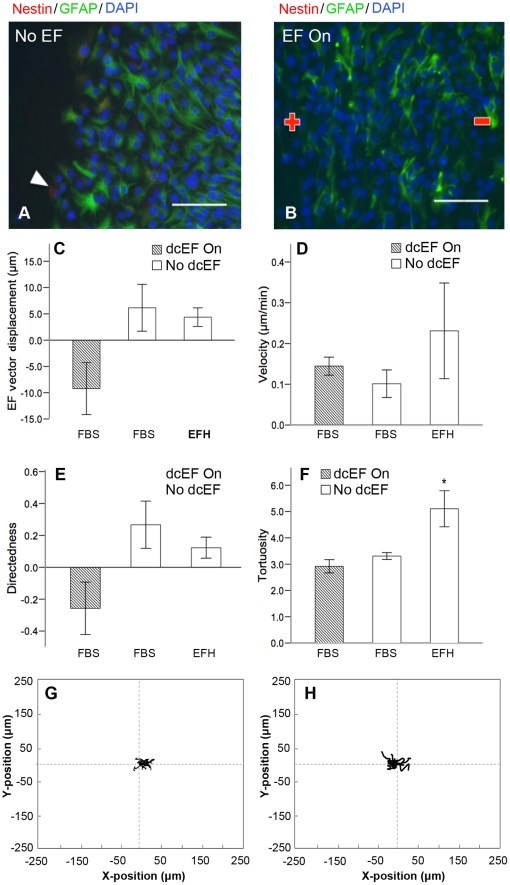
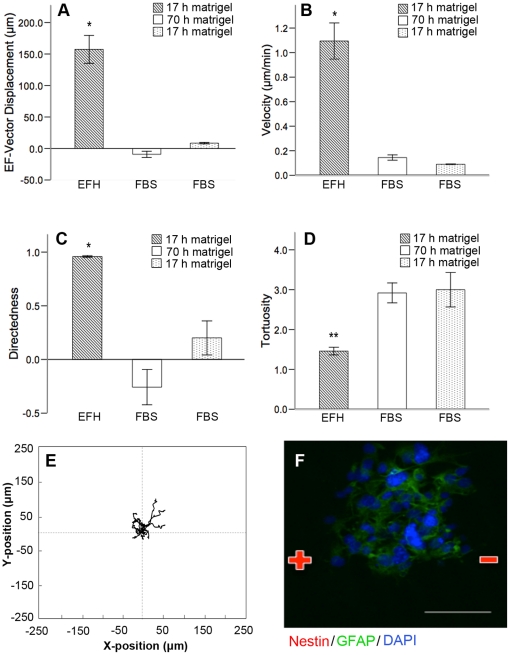
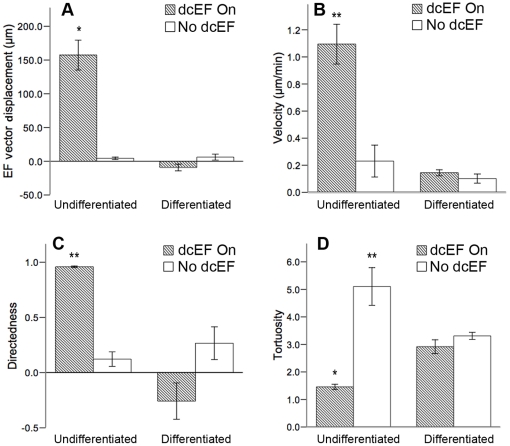
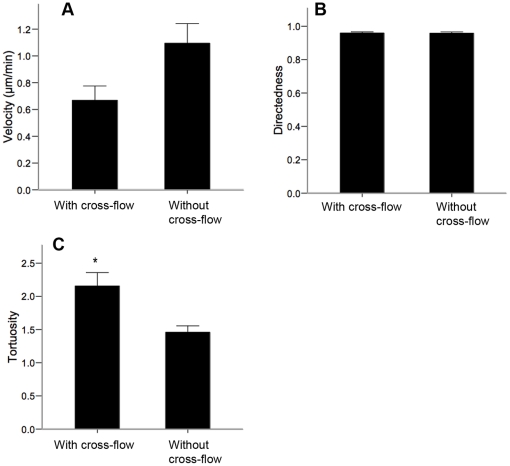
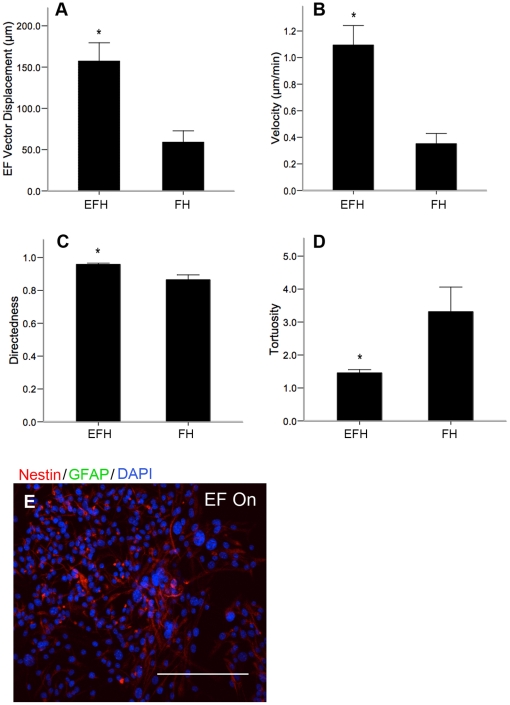
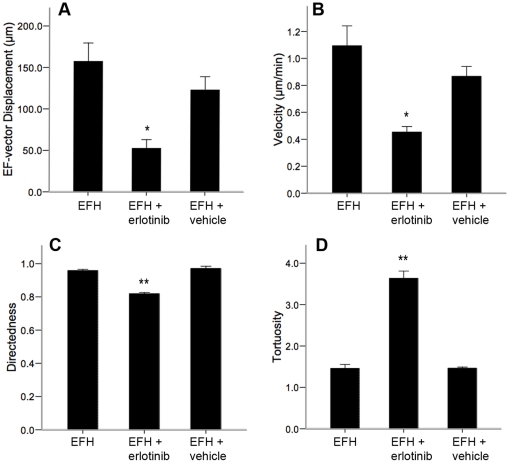
Methods and findings: With the goal of enhancing neural precursor migration to facilitate the repair process we report that externally applied direct current electric fields induce rapid and directed cathodal migration of pure populations of undifferentiated adult subependyma-derived neural precursors. Using time-lapse imaging microscopy in vitro we performed an extensive single-cell kinematic analysis demonstrating that this galvanotactic phenomenon is a feature of undifferentiated precursors, and not differentiated phenotypes. Moreover, we have shown that the migratory response of the neural precursors is a direct effect of the electric field and not due to chemotactic gradients. We also identified that epidermal growth factor receptor (EGFR) signaling plays a role in the galvanotactic response as blocking EGFR significantly attenuates the migratory behaviour.
Conclusions: These findings suggest direct current electric fields may be implemented in endogenous repair paradigms to promote migration and tissue repair following neurotrauma.
Conflict of interest statement
Figures
References
-
- Erlandsson A, Morshead C. Exploiting the properties of adult stem cells for the treatment of disease. Curr Opin Mol Ther. 2006;8:331–337. - PubMed
-
- Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med. 2002;8:963–970. - PubMed
-
- Kolb B, Morshead C, Gonzalez C, Kim M, Gregg C, et al. Growth factor-stimulated generation of new cortical tissue and functional recovery after stroke damage to the motor cortex of rats. J Cereb Blood Flow Metab. 2007;27:983–997. - PubMed
-
- Zhang RL, Zhang ZG, Wang L, Wang Y, Gousey A, et al. Activated neural stem cells contribute to stroke-induced neurogenesis and neuroblast migration toward the infarct boundary in adult rats. J Cereb Blood Flow Metab. 2004;24:441–448. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
