

Kinetic and thermodynamic features reveal that Escherichia coli BCP is an unusually versatile peroxiredoxin
- PMID: 21910476
- PMCID: PMC3204386
- DOI: 10.1021/bi200935d
Kinetic and thermodynamic features reveal that Escherichia coli BCP is an unusually versatile peroxiredoxin
Abstract
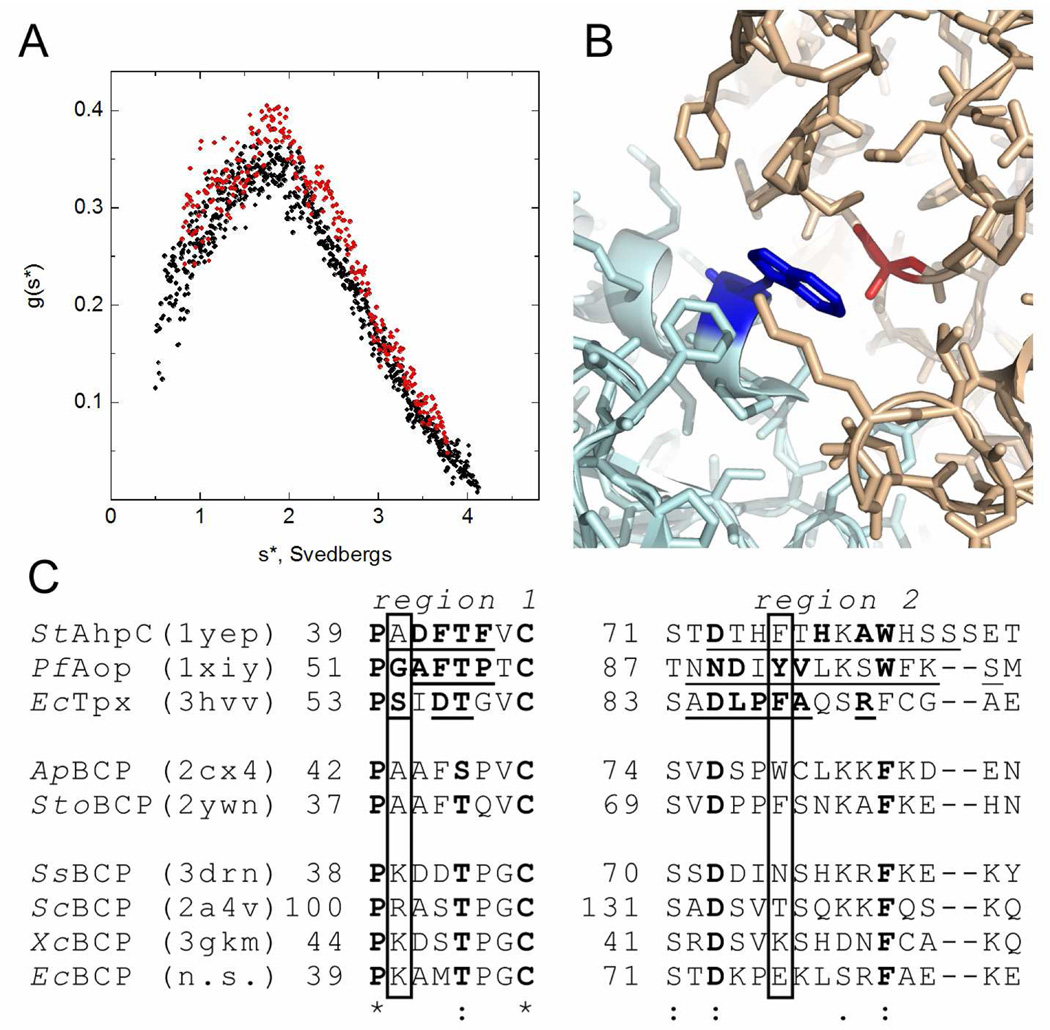
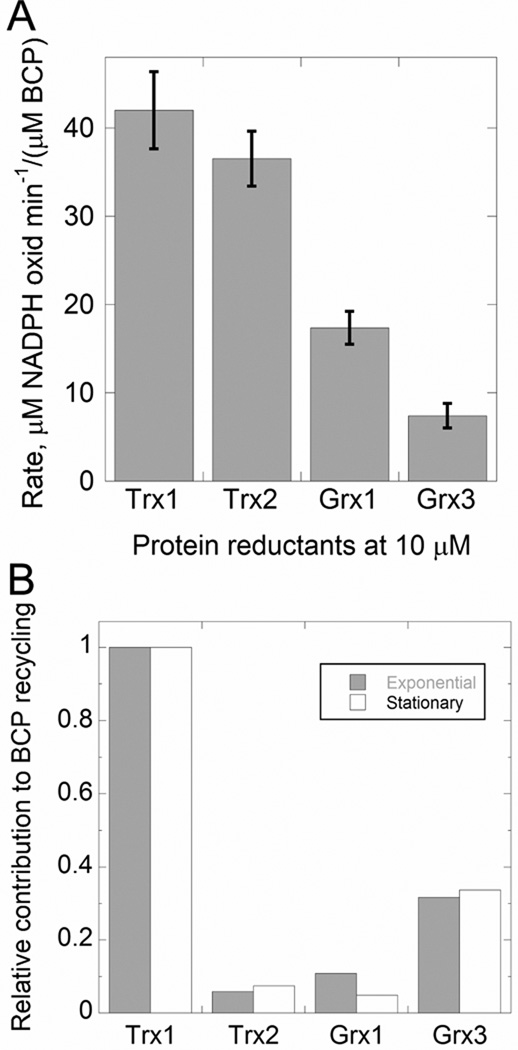
In Escherichia coli, bacterioferritin comigratory protein (BCP) is a peroxiredoxin (Prx) that catalyzes the reduction of H(2)O(2) and organic hydroperoxides. This protein, along with plant PrxQ, is a founding member of one of the least studied subfamilies of Prxs. Recent structural data have suggested that proteins in the BCP/PrxQ group can exist as monomers or dimers; we report here that, by analytical ultracentrifugation, both oxidized and reduced E. coli BCP behave as monomers in solution at concentrations as high as 200 μM. Unexpectedly, thioredoxin (Trx1)-dependent peroxidase assays conducted by stopped-flow spectroscopy demonstrated that V(max,app) increases with increasing Trx1 concentrations, indicating a nonsaturable interaction (K(m) > 100 μM). At a physiologically reasonable Trx1 concentration of 10 μM, the apparent K(m) value for H(2)O(2) is ~80 μM, and overall, the V(max)/K(m) for H(2)O(2), which remains constant at the various Trx1 concentrations (consistent with a ping-pong mechanism), is ~1.3 × 10(4) M(-1) s(-1). Our kinetic analyses demonstrated that BCP can utilize a variety of reducing substrates, including Trx1, Trx2, Grx1, and Grx3. BCP exhibited a high redox potential of -145.9 ± 3.2 mV, the highest to date observed for a Prx. Moreover, BCP exhibited a broad peroxide specificity, with comparable rates for H(2)O(2) and cumene hydroperoxide. We determined a pK(a) of ~5.8 for the peroxidatic cysteine (Cys45) using both spectroscopic and activity titration data. These findings support an important role for BCP in interacting with multiple substrates and remaining active under highly oxidizing cellular conditions, potentially serving as a defense enzyme of last resort.
© 2011 American Chemical Society
Figures



References
-
- Ryan CS, Kleinberg I. Bacteria in human mouths involved in the production and utilization of hydrogen peroxide. Arch Oral Biol. 1995;40:753–763. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
