

Myosin XIK is a major player in cytoplasm dynamics and is regulated by two amino acids in its tail
- PMID: 21914656
- PMCID: PMC3245463
- DOI: 10.1093/jxb/err265
Myosin XIK is a major player in cytoplasm dynamics and is regulated by two amino acids in its tail
Abstract
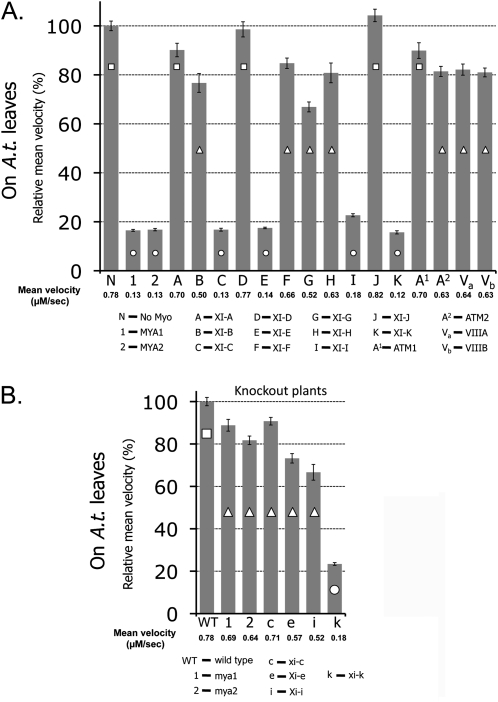
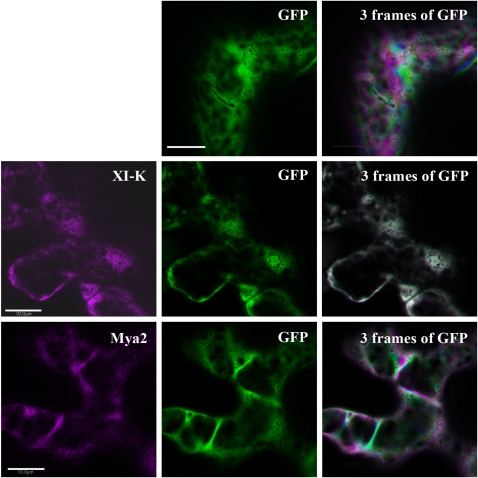
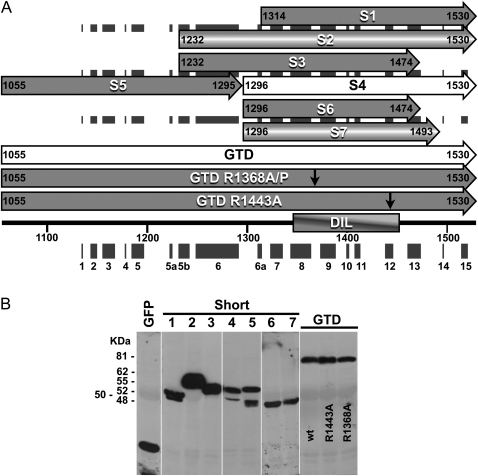
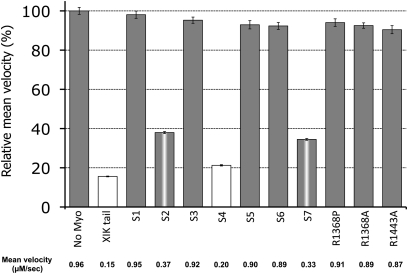
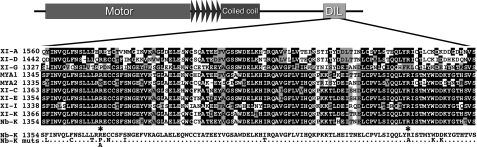
It has recently been found that among the 17 Arabidopsis myosins, six (XIC, XIE, XIK, XI-I, MYA1, and MYA2) have a major role in the motility of Golgi bodies and mitochondria in Nicotiana benthamiana and Nicotiana tabacum. Here, the same dominant negative tail fragments were also found to arrest the movement of Gogi bodies when transiently expressed in Arabidopsis plants. However, when a Golgi marker was transiently expressed in plants knocked out in these myosins, its movement was dramatically inhibited only in the xik mutant. In addition, a tail fragment of myosin XIK could inhibit the movement of several post-Golgi organelles, such as the trans-Golgi network, pre-vacuolar compartment, and endosomes, as well as total cytoplasmic streaming, suggesting that myosin XIK is a major player in cytoplasm kinetics. However, no co-localization of myosin tails with the arrested organelles was observed. Several deletion truncations of the myosin XIK tail were generated to corroborate function with localization. All deletion mutants possessing an inhibitory effect on organelle movement exhibited a diffuse cytoplasmic distribution. Point mutations in the tail of myosin XIK revealed that Arg1368 and Arg1443 are essential for its activity. These residues correspond to Lys1706 and Lys1779 from mouse myosin Va, which mediate the inhibitory head-tail interaction in this myosin. Therefore, such an interaction might underlie the dominant negative effect of truncated plant myosin tails and explain the mislocalization with target organelles.
Figures

References
-
- Arnold K, Bordoli L, Kopp J, Schwede T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics. 2006;22:195–201. - PubMed
-
- Esseling-Ozdoba A, Houtman D, van Lammeren AAM, Eiser E, Emons AM. Hydrodynamic flow in the cytoplasm of plant cells. Journal of Microscopy. 2008;231:274–283. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
