

Structure of Streptomyces maltosyltransferase GlgE, a homologue of a genetically validated anti-tuberculosis target
- PMID: 21914799
- PMCID: PMC3207445
- DOI: 10.1074/jbc.M111.279315
Structure of Streptomyces maltosyltransferase GlgE, a homologue of a genetically validated anti-tuberculosis target
Abstract
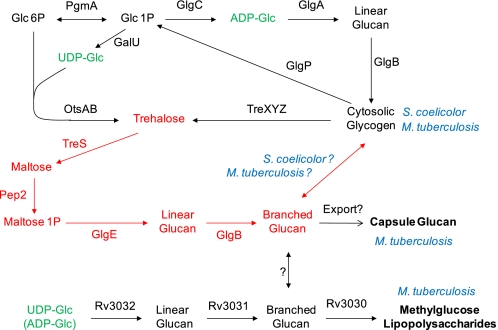
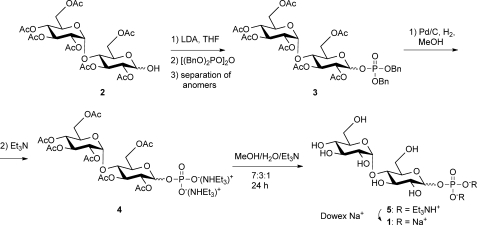
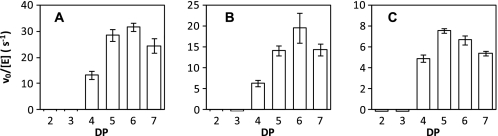
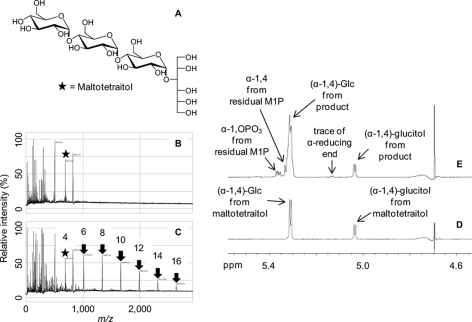
GlgE is a recently identified (1→4)-α-d-glucan:phosphate α-d-maltosyltransferase involved in α-glucan biosynthesis in bacteria and is a genetically validated anti-tuberculosis drug target. It is a member of the GH13_3 CAZy subfamily for which no structures were previously known. We have solved the structure of GlgE isoform I from Streptomyces coelicolor and shown that this enzyme has the same catalytic and very similar kinetic properties to GlgE from Mycobacterium tuberculosis. The S. coelicolor enzyme forms a homodimer with each subunit comprising five domains, including a core catalytic α-amylase-type domain A with a (β/α)(8) fold. This domain is elaborated with domain B and two inserts that are specifically configured to define a well conserved donor pocket capable of binding maltose. Domain A, together with domain N from the neighboring subunit, forms a hydrophobic patch that is close to the maltose-binding site and capable of binding cyclodextrins. Cyclodextrins competitively inhibit the binding of maltooligosaccharides to the S. coelicolor enzyme, showing that the hydrophobic patch overlaps with the acceptor binding site. This patch is incompletely conserved in the M. tuberculosis enzyme such that cyclodextrins do not inhibit this enzyme, despite acceptor length specificity being conserved. The crystal structure reveals two further domains, C and S, the latter being a helix bundle not previously reported in GH13 members. The structure provides a framework for understanding how GlgE functions and will help guide the development of inhibitors with therapeutic potential.
Figures
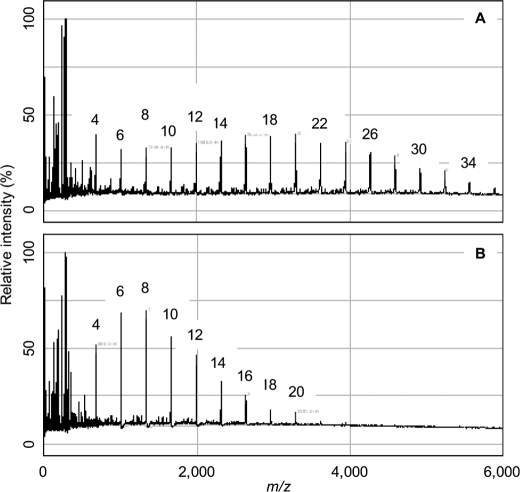
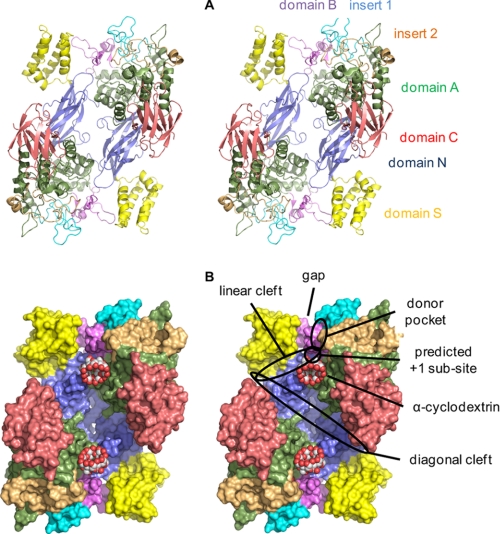
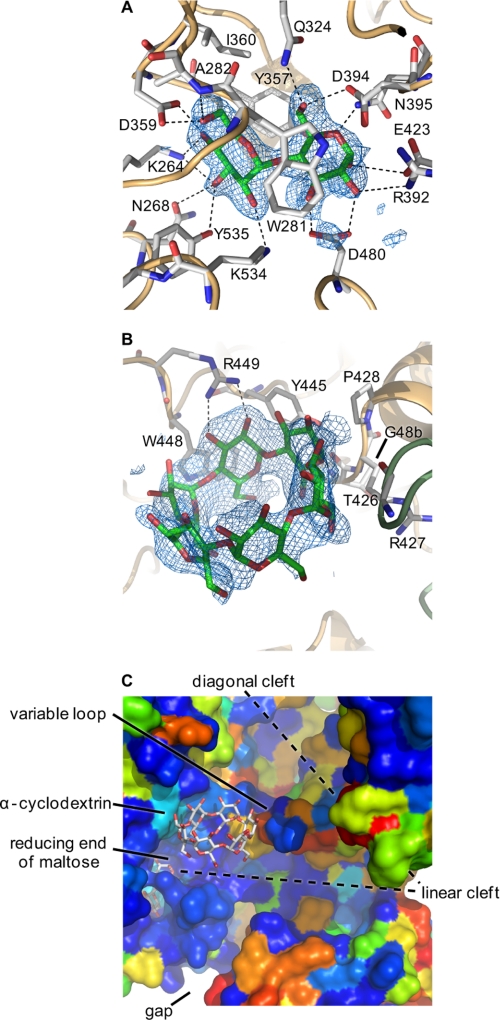
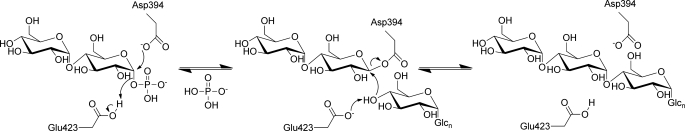
References
-
- Koul A., Arnoult E., Lounis N., Guillemont J., Andries K. (2011) Nature 469, 483–490 - PubMed
-
- Dye C. (2006) Lancet 367, 938–940 - PubMed
-
- Stam M. R., Danchin E. G., Rancurel C., Coutinho P. M., Henrissat B. (2006) Protein Eng. Des. Sel. 19, 555–562 - PubMed
-
- Kalscheuer R., Jacobs W. R., Jr. (2010) Drug News Perspect. 23, 619–624 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
