

Cardiac remodeling in fish: strategies to maintain heart function during temperature Change
- PMID: 21915331
- PMCID: PMC3168507
- DOI: 10.1371/journal.pone.0024464
Cardiac remodeling in fish: strategies to maintain heart function during temperature Change
Abstract
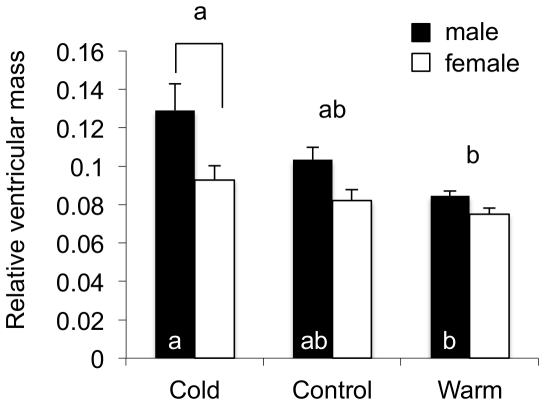
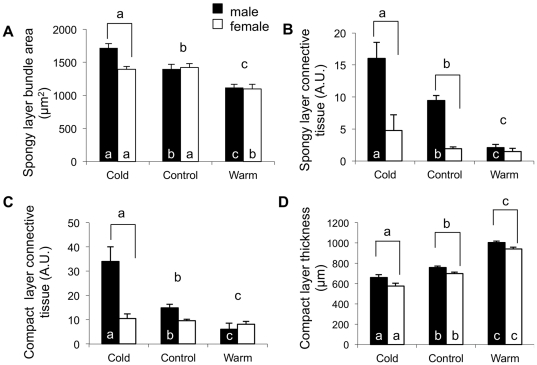
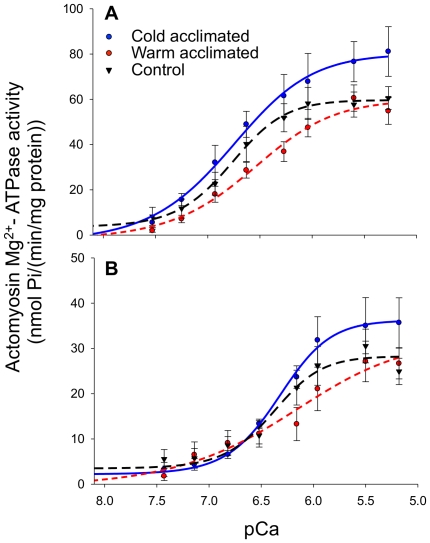
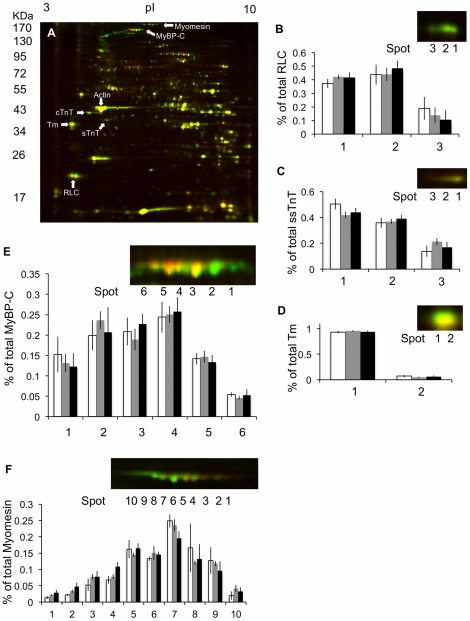
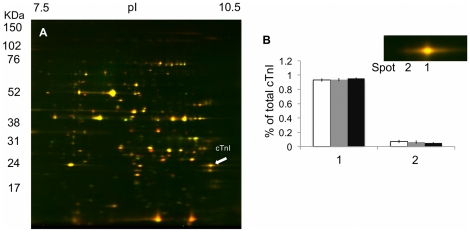
Rainbow trout remain active in waters that seasonally change between 4°C and 20°C. To explore how these fish are able to maintain cardiac function over this temperature range we characterized changes in cardiac morphology, contractile function, and the expression of contractile proteins in trout following acclimation to 4°C (cold), 12°C (control), and 17°C (warm). The relative ventricular mass (RVM) of the cold acclimated male fish was significantly greater than that of males in the control group. In addition, the compact myocardium of the cold acclimated male hearts was thinner compared to controls while the amount of spongy myocardium was found to have increased. Cold acclimation also caused an increase in connective tissue content, as well as muscle bundle area in the spongy myocardium of the male fish. Conversely, warm acclimation of male fish caused an increase in the thickness of the compact myocardium and a decrease in the amount of spongy myocardium. There was also a decrease in connective tissue content in both myocardial layers. In contrast, there was no change in the RVM or connective tissue content in the hearts of female trout with warm or cold acclimation. Cold acclimation also caused a 50% increase in the maximal rate of cardiac AM Mg(2+)-ATPase but did not influence the Ca(2+) sensitivity of this enzyme. To identify a mechanism for this change we utilized two-dimensional difference gel electrophoresis to characterize changes in the cardiac contractile proteins. Cold acclimation caused subtle changes in the phosphorylation state of the slow skeletal isoform of troponin T found in the heart, as well as of myosin binding protein C. These results demonstrate that acclimation of trout to warm and cold temperatures has opposing effects on cardiac morphology and tissue composition and that this results in distinct warm and cold cardiac phenotypes.
Conflict of interest statement
Figures
References
-
- Bernardo BC, Weeks KL, Pretorius L, McMullen JR. Molecular distinction between physiological and pathological cardiac hypertrophy: Experimental findings and therapeutic strategies. Pharm. 2010;Ther128:191–227. - PubMed
-
- McMullen JR, Jennings GL. Differences between pathological and physiological cardiac hypertrophy: Novel therapeutic strategies to treat heart failure. Clin Exp Pharmacol Physiol. 2007;34:255–262. - PubMed
-
- Rupp H. The adaptive changes in the isoenzyme pattern of myosin from hypertrophied rat myocardium as a result of pressure overload and physical training. Basic Res Cardiol. 1981;76:79–88. - PubMed
-
- Wikman-Coffelt J, Parmley WW, Mason DT. The cardiac hypertrophy process. Analyses of factors determining pathological vs. physiological development. Circ Res. 1979;45:697–707. - PubMed
-
- Farrell AP, Hammons AM, Graham MS, Tibbits GF. Cardiac growth in rainbow trout, Salmo gairdneri. Can J Zool. 1988;66:2368–73.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
