

Molecular characterization of a strawberry FaASR gene in relation to fruit ripening
- PMID: 21915355
- PMCID: PMC3167850
- DOI: 10.1371/journal.pone.0024649
Molecular characterization of a strawberry FaASR gene in relation to fruit ripening
Abstract
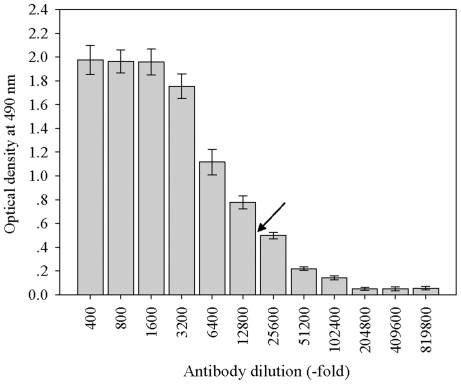
Background: ABA-, stress- and ripening-induced (ASR) proteins have been reported to act as a downstream component involved in ABA signal transduction. Although much attention has been paid to the roles of ASR in plant development and stress responses, the mechanisms by which ABA regulate fruit ripening at the molecular level are not fully understood. In the present work, a strawberry ASR gene was isolated and characterized (FaASR), and a polyclonal antibody against FaASR protein was prepared. Furthermore, the effects of ABA, applied to two different developmental stages of strawberry, on fruit ripening and the expression of FaASR at transcriptional and translational levels were investigated.
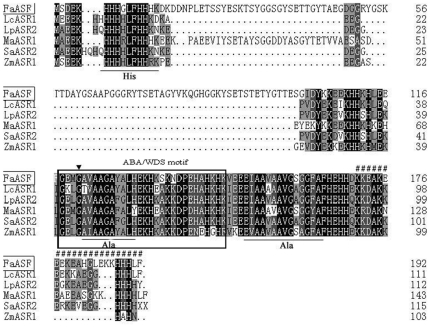
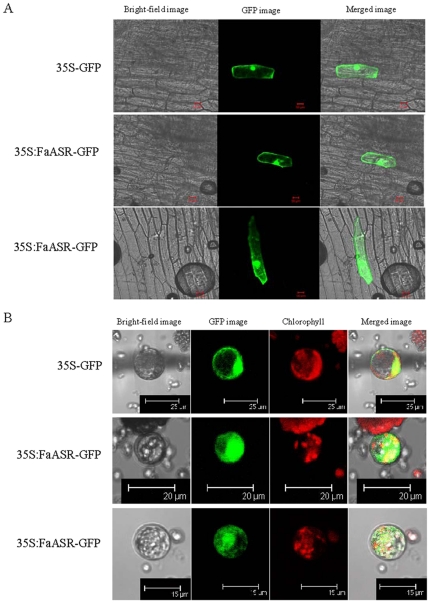
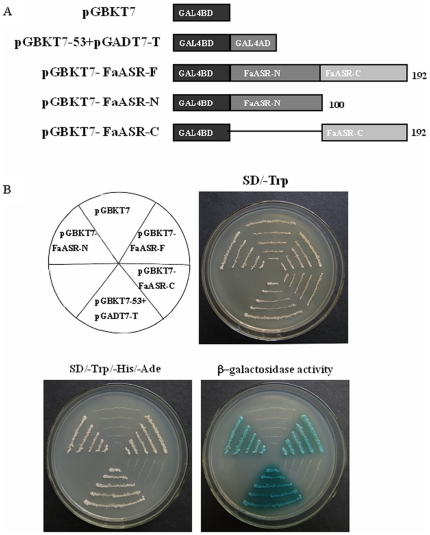
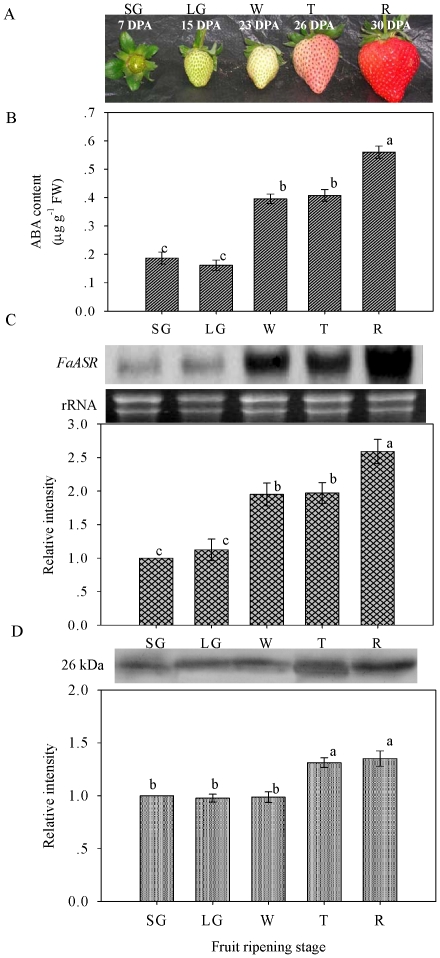
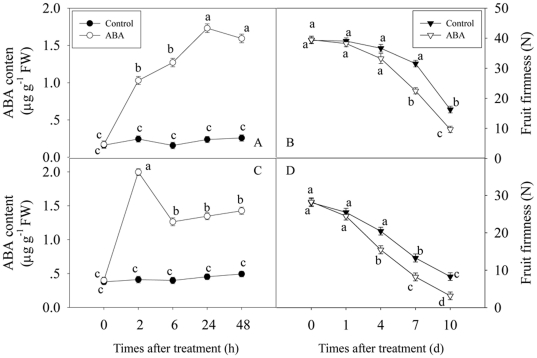
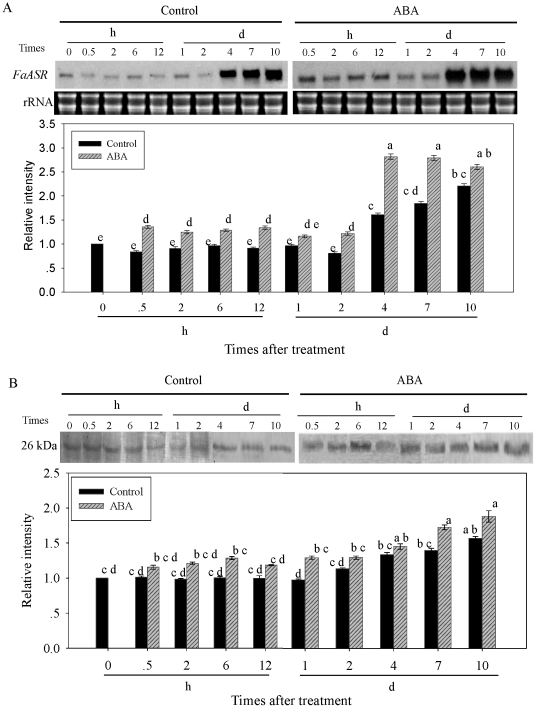
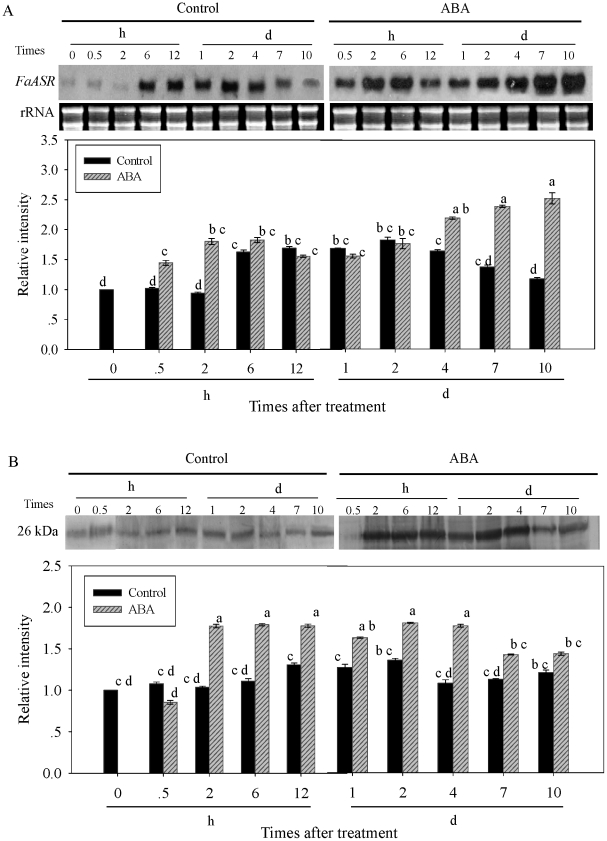
Methodology/principal findings: FaASR, localized in the cytoplasm and nucleus, contained 193 amino acids and shared common features with other plant ASRs. It also functioned as a transcriptional activator in yeast with trans-activation activity in the N-terminus. During strawberry fruit development, endogenous ABA content, levels of FaASR mRNA and protein increased significantly at the initiation of ripening at a white (W) fruit developmental stage. More importantly, application of exogenous ABA to large green (LG) fruit and W fruit markedly increased endogenous ABA content, accelerated fruit ripening, and greatly enhanced the expression of FaASR transcripts and the accumulation of FaASR protein simultaneously.
Conclusions: These results indicate that FaASR may be involved in strawberry fruit ripening. The observed increase in endogenous ABA content, and enhanced FaASR expression at transcriptional and translational levels in response to ABA treatment might partially contribute to the acceleration of strawberry fruit ripening.
Conflict of interest statement
Figures

References
-
- Giovannoni JJ. Molecular biology of fruit maturation and ripening. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:725–749. - PubMed
-
- Alexander L, Grierson D. Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening. J Exp Bot. 2002;53:2039–2055. - PubMed
-
- Manning K. Isolation of a set of ripening-related genes from strawberry: their identification and possible relationship to fruit quality traits. Planta. 1998;205:622–631. - PubMed
-
- Chervin C, El-Kereamy A, Roustan JP, Latché A, Lamon J, et al. Ethylene seems required for the berry development and ripening in grape, a non-climacteric fruit. Plant Sci. 2004;167:1301–1305.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
