

Celiac disease and transglutaminase 2: a model for posttranslational modification of antigens and HLA association in the pathogenesis of autoimmune disorders
- PMID: 21917438
- PMCID: PMC3428143
- DOI: 10.1016/j.coi.2011.08.006
Celiac disease and transglutaminase 2: a model for posttranslational modification of antigens and HLA association in the pathogenesis of autoimmune disorders
Abstract
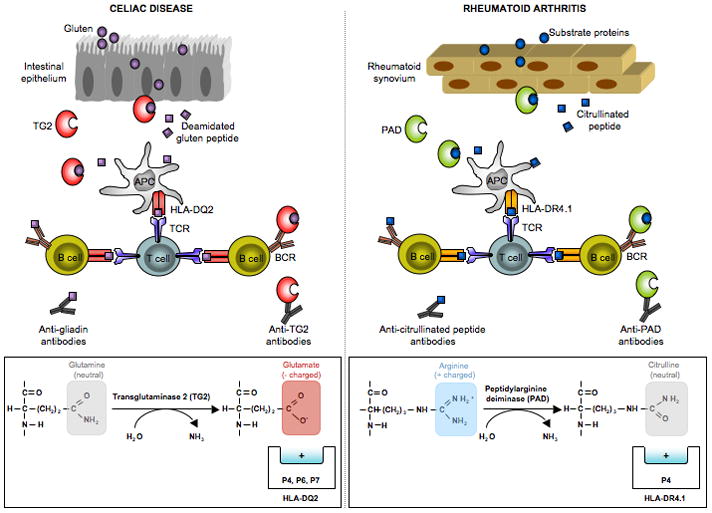
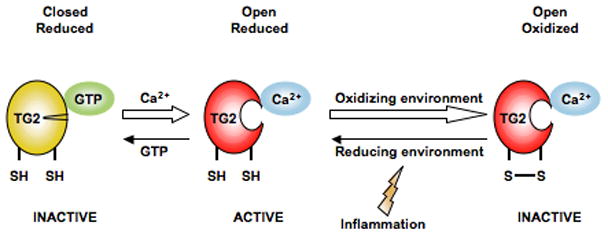
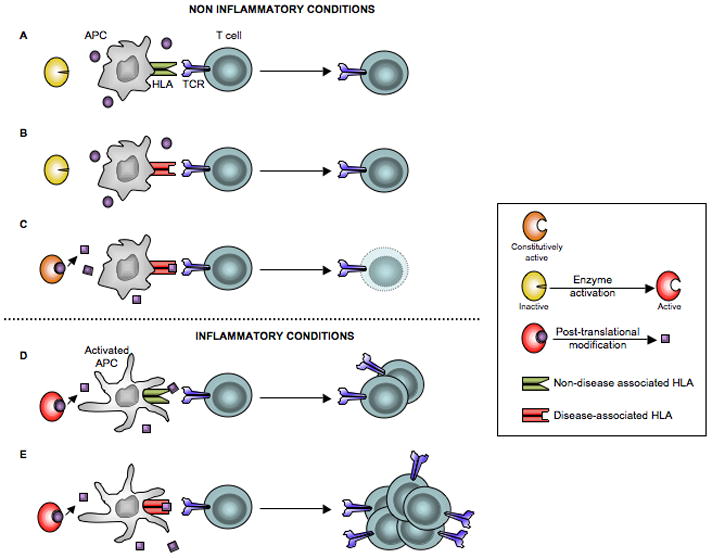
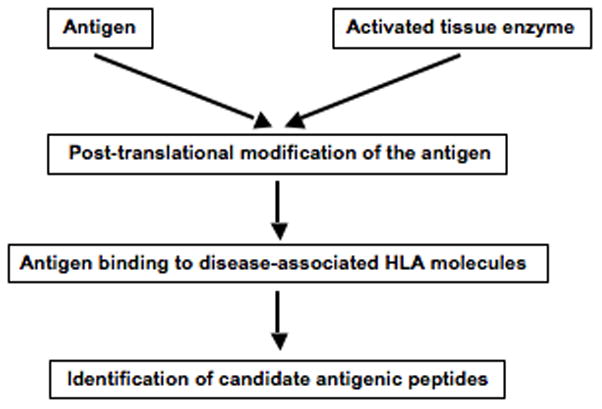
Posttranslational modification (PTM) of antigen is a way to break T-cell tolerance to self-antigens and promote autoimmunity. However, the precise mechanisms by which modifications would facilitate autoimmune T-cell responses and how they relate to particular autoimmune-associated MHC molecules remain elusive. Celiac disease is a T-cell mediated enteropathy with a strong HLA association where the immune response is directed mainly against deamidated cereal gluten peptides that have been modified by the enzyme transglutaminase 2. The disease is further characterized by autoantibodies to transglutaminase 2 that have extraordinary high disease specificity and sensitivity. There have been important advances in the knowledge of celiac disease pathogenesis, and these insights may be applicable to other autoimmune disorders where PTM plays a role. This insight gives clues for understanding the involvement of PTMs in other autoimmune diseases.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Trynka G, Wijmenga C, van Heel DA. A genetic perspective on coeliac disease. Trends Mol Med. 2010;16:537–550. - PubMed
-
- Karell K, Louka AS, Moodie SJ, Ascher H, Clot F, Greco L, Ciclitira PJ, Sollid LM, Partanen J. HLA types in celiac disease patients not carrying the DQA1*05-DQB1*02 (DQ2) heterodimer: results from the European Genetics Cluster on Celiac Disease. Hum Immunol. 2003;64:469–477. - PubMed
-
- Dubois PC, Trynka G, Franke L, Hunt KA, Romanos J, Curtotti A, Zhernakova A, Heap GA, Adany R, Aromaa A, et al. Multiple common variants for celiac disease influencing immune gene expression. Nat Genet. 2010;42:295–302. The most comprensive genome wide association study of celiac disease identifying 39 non-HLA susceptibility loci. - PMC - PubMed
-
- Lorand L, Graham RM. Transglutaminases: crosslinking enzymes with pleiotropic functions. Nat Rev Mol Cell Biol. 2003;4:140–156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
