

Gene length may contribute to graded transcriptional responses in the Drosophila embryo
- PMID: 21920356
- PMCID: PMC3661008
- DOI: 10.1016/j.ydbio.2011.08.016
Gene length may contribute to graded transcriptional responses in the Drosophila embryo
Abstract
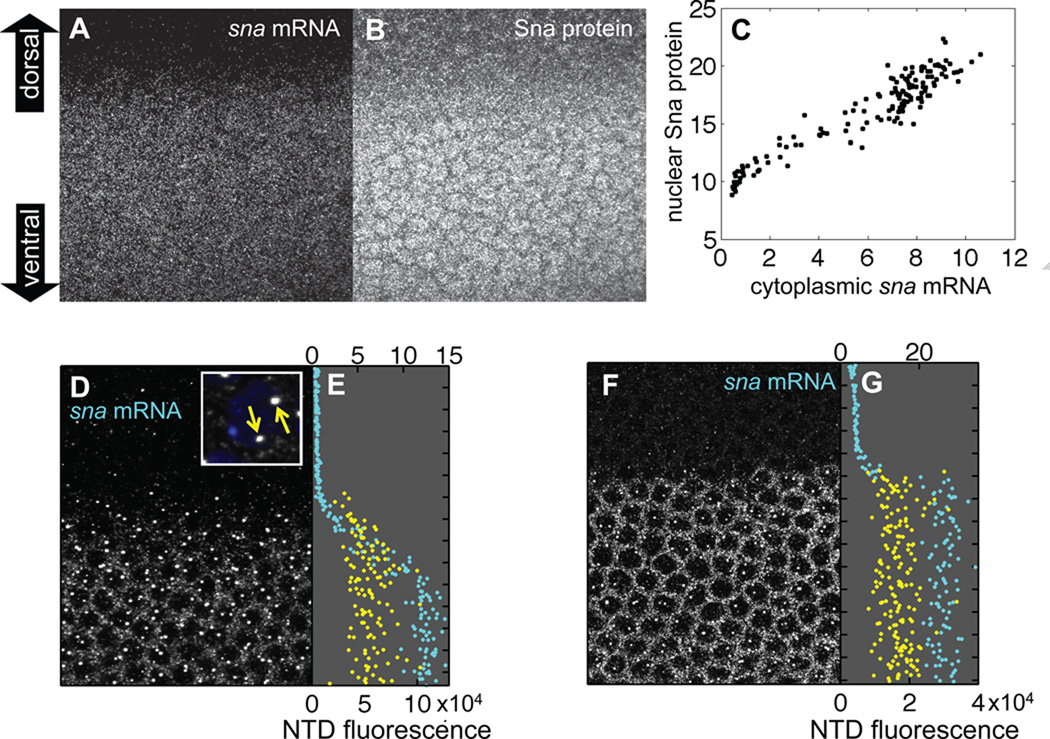
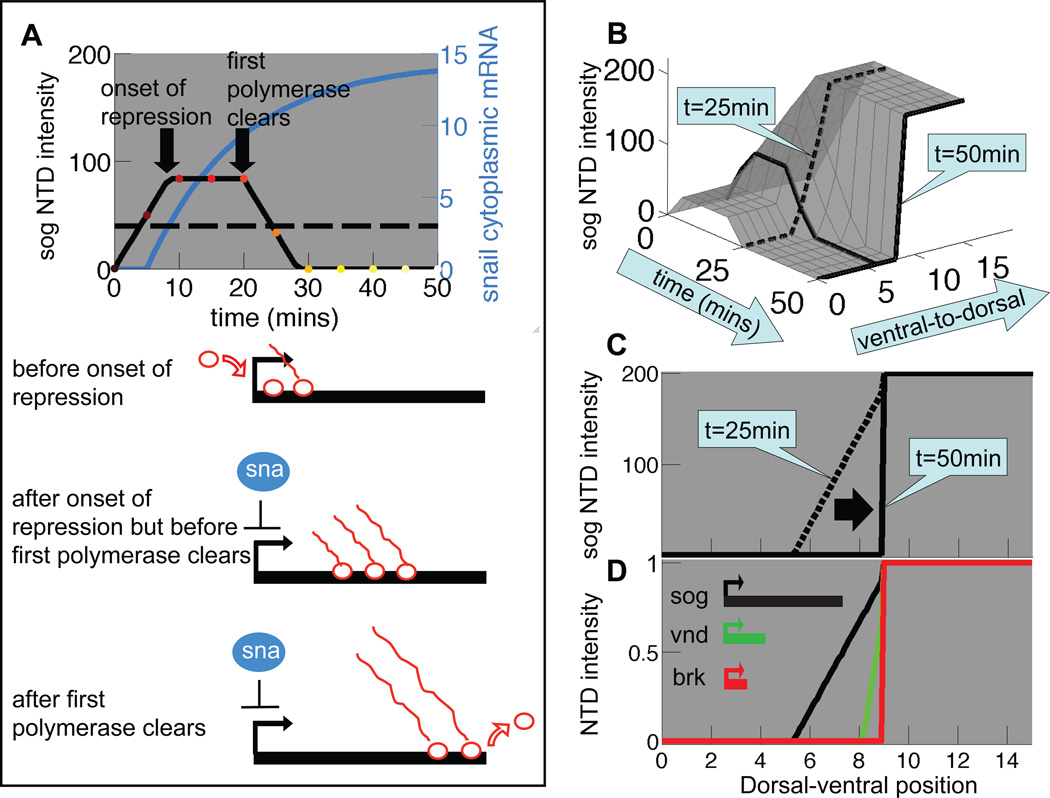
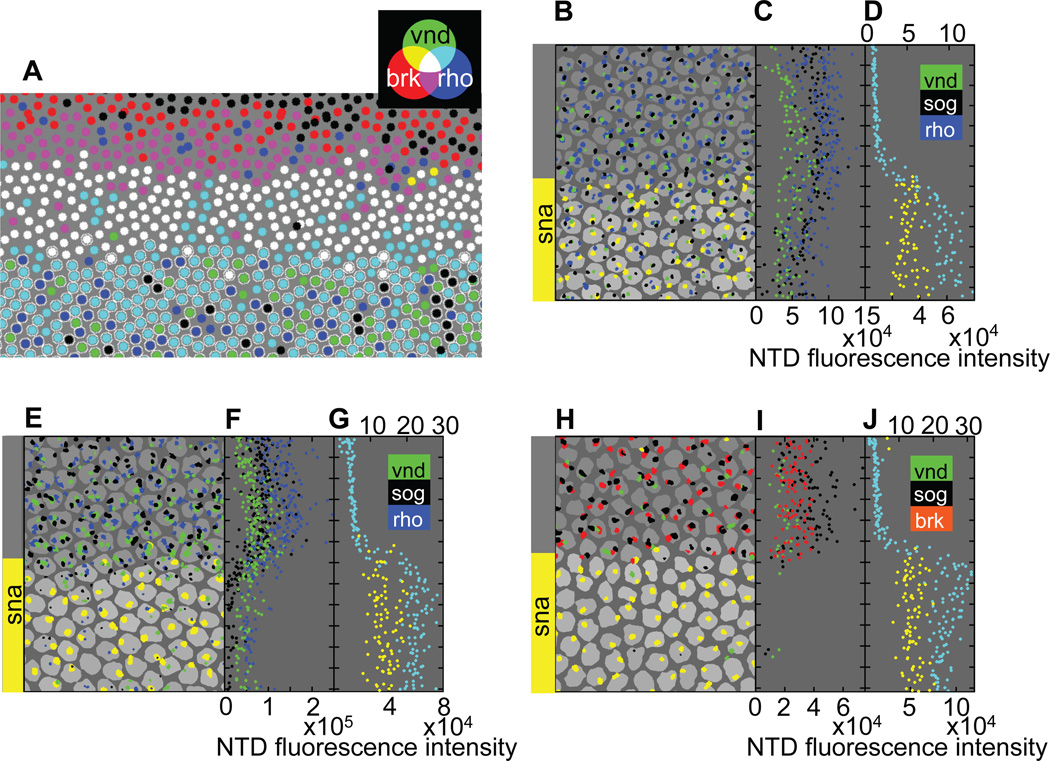
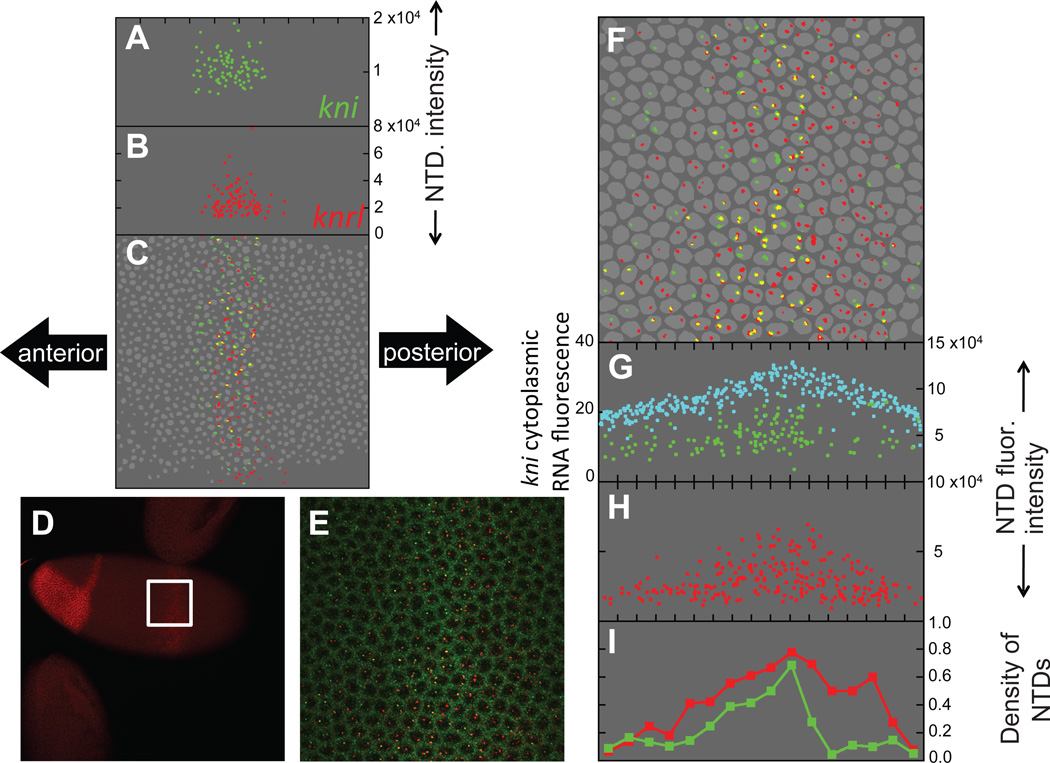
An important question in developmental biology is how relatively shallow gradients of morphogens can reliably establish a series of distinct transcriptional readouts. Current models emphasize interactions between transcription factors binding in distinct modes to cis-acting sequences of target genes. Another recent idea is that the cis-acting interactions may amplify preexisting biases or prepatterns to establish robust transcriptional responses. In this study, we examine the possible contribution of one such source of prepattern, namely gene length. We developed quantitative imaging tools to measure gene expression levels for several loci at a time on a single-cell basis and applied these quantitative imaging tools to dissect the establishment of a gene expression border separating the mesoderm and neuroectoderm in the early Drosophila embryo. We first characterized the formation of a transient ventral-to-dorsal gradient of the Snail (Sna) repressor and then examined the relationship between this gradient and repression of neural target genes in the mesoderm. We found that neural genes are repressed in a nested pattern within a zone of the mesoderm abutting the neuroectoderm, where Sna levels are graded. While several factors may contribute to the transient graded response to the Sna gradient, our analysis suggests that gene length may play an important, albeit transient, role in establishing these distinct transcriptional responses. One prediction of the gene-length-dependent transcriptional patterning model is that the co-regulated genes knirps (a short gene) and knirps-related (a long gene) should be transiently expressed in domains of differing widths, which we confirmed experimentally. These findings suggest that gene length may contribute to establishing graded responses to morphogen gradients by providing transient prepatterns that are subsequently amplified and stabilized by traditional cis-regulatory interactions.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


References
-
- Bardin AJ, Schweisguth F. Bearded family members inhibit Neuralized-mediated endocytosis and signaling activity of Delta in Drosophila. Dev Cell. 2006;10:245–255. - PubMed
-
- Barolo S, Posakony JW. Three habits of highly effective signaling pathways: principles of transcriptional control by developmental cell signaling. Genes Dev. 2002;16:1167–1181. - PubMed
-
- Capovilla M, Eldon E, Pirrotta V. The giant gene of Drosophila encodes a B-ZIP DNA-binding protein that regulates the expression of other segmentation gap genes. Development. 1992;114:99–112. - PubMed
-
- Chitnis AB. Keeping single minded expression on the straight and narrow. Mol Cell. 2006;21:450–452. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
