

Effect of the ILE86TER mutation in the γ subunit of cGMP phosphodiesterase (PDE6) on rod photoreceptor signaling
- PMID: 21920434
- PMCID: PMC3205198
- DOI: 10.1016/j.cellsig.2011.08.021
Effect of the ILE86TER mutation in the γ subunit of cGMP phosphodiesterase (PDE6) on rod photoreceptor signaling
Abstract
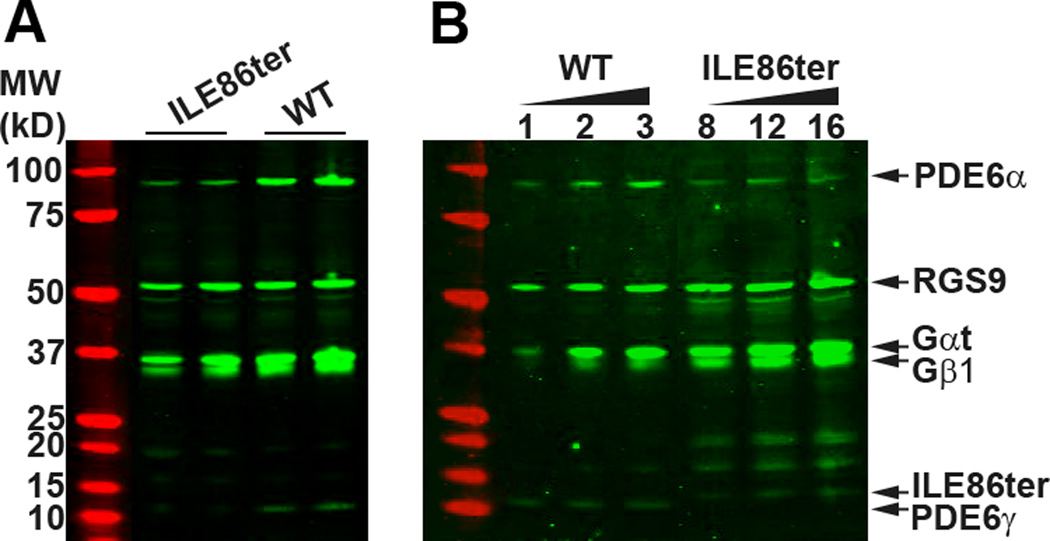
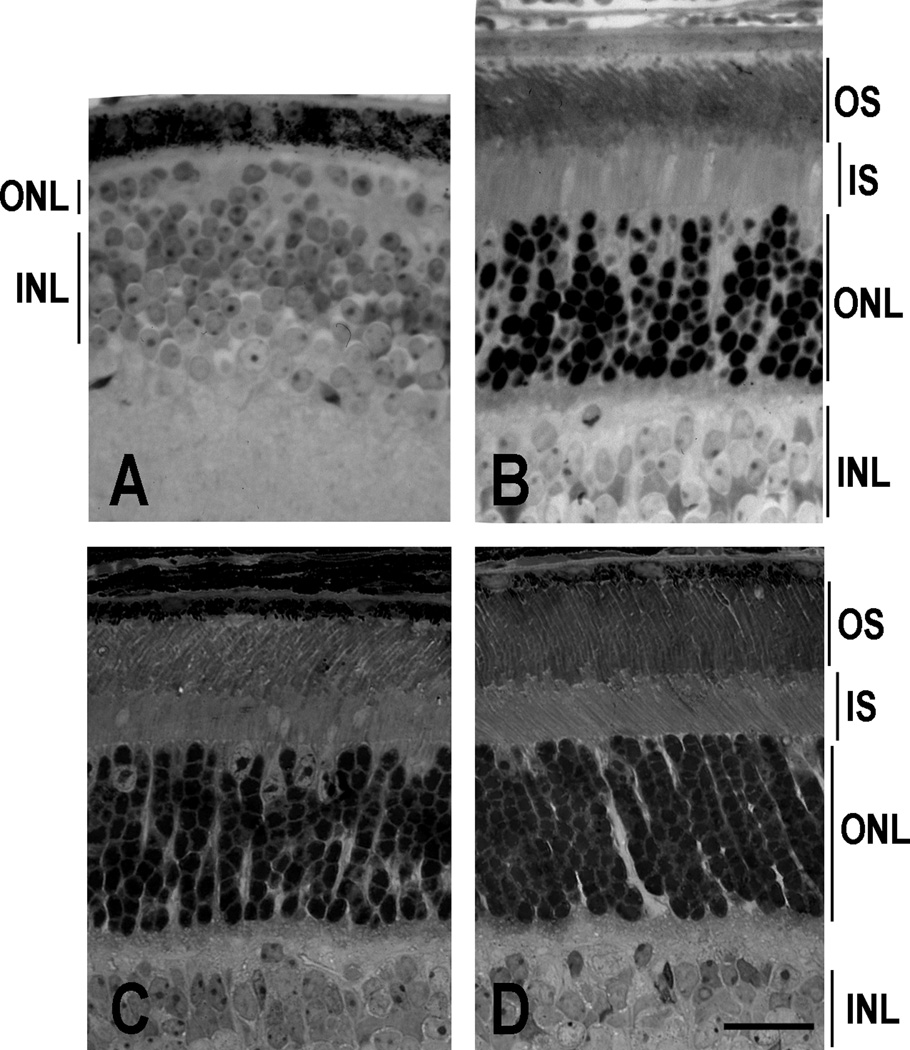
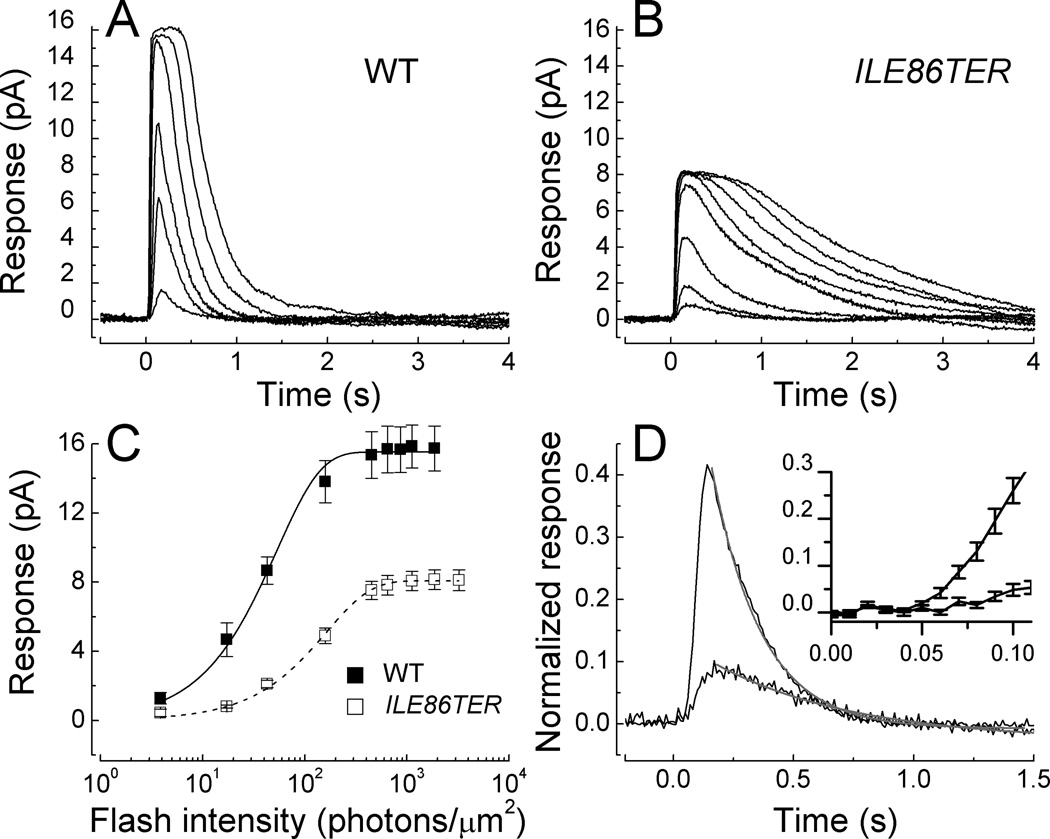
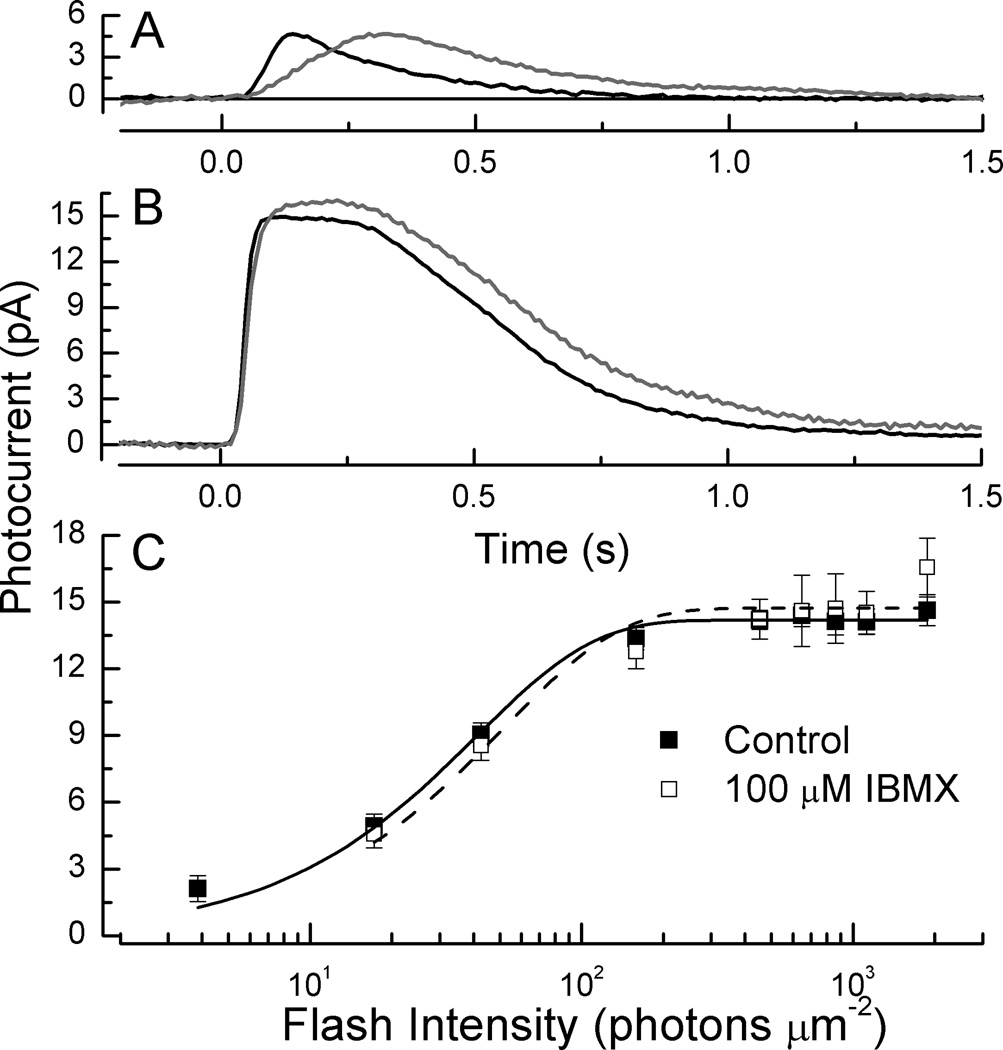
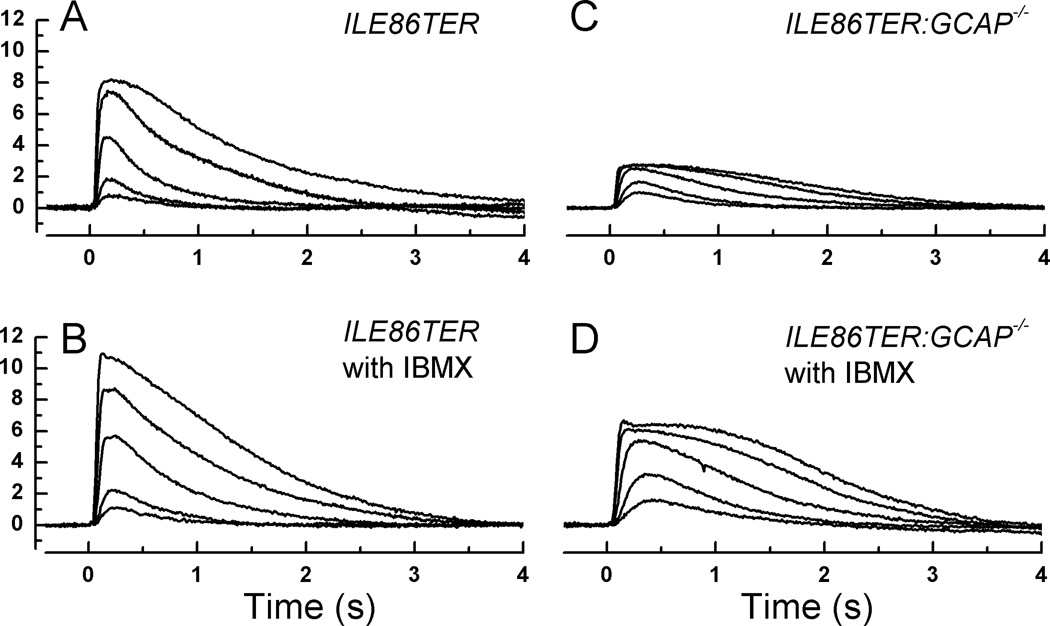
The light-dependent decrease in cyclic guanosine monophosphate (cGMP) in the rod outer segment is produced by a phosphodiesterase (PDE6), consisting of catalytic α and β subunits and two inhibitory γ subunits. The molecular mechanism of PDE6γ regulation of the catalytic subunits is uncertain. To study this mechanism in vivo, we introduced a modified Pde6g gene for PDE6γ into a line of Pde6g(tm1)/Pde6g(tm1) mice that do not express PDE6γ. The resulting ILE86TER mice have a PDE6γ that lacks the two final carboxyl-terminal Ile(86) and Ile(87) residues, a mutation previously shown in vitro to reduce inhibition by PDE6γ. ILE86TER rods showed a decreased sensitivity and rate of activation, probably the result of a decreased level of expression of PDE6 in ILE86TER rods. More importantly, they showed a decreased rate of decay of the photoresponse, consistent with decreased inhibition of PDE6 α and β by PDE6γ. Furthermore, ILE86TER rods had a higher rate of spontaneous activation of PDE6 than WT rods. Circulating current in ILE86TER rods that also lacked both guanylyl cyclase activating proteins (GCAPs) could be increased several fold by perfusion with 100μM of the PDE6 inhibitor 3-isobutyl-1-methylxanthine (IBMX), consistent with a higher rate of dark PDE6 activity in the mutant photoreceptors. In contrast, IBMX had little effect on the circulating current of WT rods, unlike previous results from amphibians. Our results show for the first time that the Ile(86) and Ile(87) residues are necessary for normal inhibition of PDE6 catalytic activity in vivo, and that increased basal activity of PDE can be partially compensated by GCAP-dependent regulation of guanylyl cyclase.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Fain GL. Sensory Transduction. Sunderland, MA: Sinauer, Inc.; 2003.
-
- Granovsky AE, Artemyev NO. Biochemistry. 2001;40(44):13209–13215. - PubMed
-
- Lipkin VM, Dumler IL, Muradov KG, Artemyev NO, Etingof RN. FEBS Lett. 1988;234(2):287–290. - PubMed
-
- Gonzalez K, Cunnick J, Takemoto D. Biochem Biophys Res Commun. 1991;181(3):1094–1096. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
